| Login | Einklappen |
Startseite
Artbeschreibungen
Allobates
Anomaloglossus
Rheobates
Aromobates
Mannophryne
Ameerega
Colostethus
Epipedobates
Silverstoneia
Andinobates
Adelphobates
Dendrobates
Excidobates
Minyobates
Oophaga
Phyllobates
Ranitomeya
Hyloxalus
Foren
Inserate
Reportagen
Service & Links
Einstellungen
| Fotos von Nutzern | Einklappen |
| Nomenklatur | Einklappen |
wissenschaftl. Name: Oophaga vicentei (JUNGFER, WEYGOLDT & JURASKE, 1996)
Etymologie:
Synonyme:
Oophaga vicentei (GRANT, FROST, CALDWELL, GAGLIARDO, HADDAD, KOK, MEANS, NOONAN, SCHARGEL & WHEELER, 2006)
Dendrobates vicentei (JUNGFER, WEYGOLD & JURASKE 1996)
sensu FROST (2006)
englischer Name: Vicent´s Poison Frog
deutscher Name: Vicentes-Baumsteiger
Etymologie:
Der Gattungsname Oophaga (fem.) leitet sich vom lateinischen oophag (= Ei liebend) ab. Er bezieht sich auf die Tatsache, dass die Larven aller Arten der Gattung in ihrer Ernährung auf speziell vom Weibchen produzierte Nähreier (Abortiveier) angewiesen sind. Das vicentei ist ein sname zu Ehren von VICENTE AUGUSTO FERNANDEZ. Dieser half bei der Suche nach den Fröschen und sammelte selbst einige Exemplare. Er kam bei einem tragischen Unfall ums Leben.
Synonyme:
Oophaga vicentei (GRANT, FROST, CALDWELL, GAGLIARDO, HADDAD, KOK, MEANS, NOONAN, SCHARGEL & WHEELER, 2006)
Dendrobates vicentei (JUNGFER, WEYGOLD & JURASKE 1996)
sensu FROST (2006)
englischer Name: Vicent´s Poison Frog
deutscher Name: Vicentes-Baumsteiger
| Systematik | Einklappen |
Klassifizierung:
Amphibia->Anura->Dendrobatoidea->Dendrobatidae->Dendrobatinae->Oophaga->Oophaga vicentei (JUNGFER, WEYGOLDT & JURASKE, 1996)
Gruppe:
Anmerkung zur Systematik:
Amphibia->Anura->Dendrobatoidea->Dendrobatidae->Dendrobatinae->Oophaga->Oophaga vicentei (JUNGFER, WEYGOLDT & JURASKE, 1996)
Gruppe:
Die Gattung Oophaga BAUER 1994 entspricht der ehemaligen Histrionicus-Gruppe im Sinne von MYERS (1984) bzw. der Einteilung von SILVERSTONE (1975) in Pumilio- und Histrionicus-Gruppe.
Anmerkung zur Systematik:
Aufgrund der Konstantheit hinsichtlich der genetischen Charakteristika (monophyletisch abgrenzbare Gruppe), der gemeinsamen Verhaltensparameter (z.B. Zirp-Ruf, weibliche Brutpflege) und morphologischen Besonderheiten (z.B. Larvenmorphologie) wurden die Arten der ehemaligen Dendrobates histrionicus-Gruppe (sensu MEYER & DALY, 1984) von GRANT ET AL. (2006) in eine eigenständige Gattung überstellt. Für diese wurde der verfügbare ältere Name Oophaga BAUER, 1994 wieder gültig (bisher Synonym zu Dendrobates WAGLER, 1830).
Ob es sich, bei den als "Rote Vicentei" bezeichneten Tieren, ebenfalls um O. vicentei oder eine unbeschriebene Art handelt bleibt abzuwarten. Neueste genetische Untersuchungen von HAGEMANN (pers. Komm.) weisen auf eine nähere Verwandschaft dieser rot-schwarz genetzten Tiere zur nördlichen Klade (O. cf. typographa, O. speciosa und O. spec. "Escudo") hin. Auch O. vicentei von der Terra typica sind näher mit dieser Klade verwandt als mit O. pumilio s.s. (siehe Stammbaum der Gruppenübersicht) Es könnte sich also auch bei O. vicentei um eine hochpolymorphe Art handeln. Bis zur Klärung durch weitere genetische Untersuchung oder vergleichende Rufanalysen bezeichnen wir die Rote Population zunächst als O. cf. vicentei.
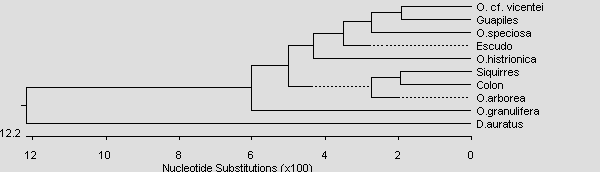
Abb.: Stammbaumartige Darstellung der Verwandtschaftsverhältnisse der Gattung Oophaga. Neighbour Joining Tree basierend auf COI-Sequenzen.
© 2007 S. Hagemann
Ob es sich, bei den als "Rote Vicentei" bezeichneten Tieren, ebenfalls um O. vicentei oder eine unbeschriebene Art handelt bleibt abzuwarten. Neueste genetische Untersuchungen von HAGEMANN (pers. Komm.) weisen auf eine nähere Verwandschaft dieser rot-schwarz genetzten Tiere zur nördlichen Klade (O. cf. typographa, O. speciosa und O. spec. "Escudo") hin. Auch O. vicentei von der Terra typica sind näher mit dieser Klade verwandt als mit O. pumilio s.s. (siehe Stammbaum der Gruppenübersicht) Es könnte sich also auch bei O. vicentei um eine hochpolymorphe Art handeln. Bis zur Klärung durch weitere genetische Untersuchung oder vergleichende Rufanalysen bezeichnen wir die Rote Population zunächst als O. cf. vicentei.
Abb.: Stammbaumartige Darstellung der Verwandtschaftsverhältnisse der Gattung Oophaga. Neighbour Joining Tree basierend auf COI-Sequenzen.
© 2007 S. Hagemann
| Bedrohungsstatus | Einklappen |
Anhang II des WA. Anhang B der EUArtschVO. Meldepflichtig nach BArtSchVO.
In der Roten Liste, wegen fehlender Freilanddaten, als nicht einstufbar bewertet. (IUCN, 2004). Laut IUCN nur aus der bekannt (El Copé). Wir konnten aber weiterhin stabile Populationen in El Valle de Antón und im P.N. Santa Fé nachweisen (OSTROWSKI, pers. Beobachtung). Alle bekannten Vorkommen liegen in Schutzgebieten und das Verbreitungsgebiet dürfte wohl noch größer sein, wie illegale Importe weiterer Varianten nach Europa 2004 bewiesen. Deshalb dürfte die Art nicht akut bedroht sein.
Achtung! Legal nach Europa eingeführten Tiere (CITES, 2005) sind Tiere von der Terra typica durch JUNGFER und Tiere von Santa Fé durch BIRKHAHN. Die einzige legale Zuchtgruppe in Deutschland dürfte die von BRANDT sein, der seine Tiere von BIRKHAHN erhielt.
In der Roten Liste, wegen fehlender Freilanddaten, als nicht einstufbar bewertet. (IUCN, 2004). Laut IUCN nur aus der bekannt (El Copé). Wir konnten aber weiterhin stabile Populationen in El Valle de Antón und im P.N. Santa Fé nachweisen (OSTROWSKI, pers. Beobachtung). Alle bekannten Vorkommen liegen in Schutzgebieten und das Verbreitungsgebiet dürfte wohl noch größer sein, wie illegale Importe weiterer Varianten nach Europa 2004 bewiesen. Deshalb dürfte die Art nicht akut bedroht sein.
Achtung! Legal nach Europa eingeführten Tiere (CITES, 2005) sind Tiere von der Terra typica durch JUNGFER und Tiere von Santa Fé durch BIRKHAHN. Die einzige legale Zuchtgruppe in Deutschland dürfte die von BRANDT sein, der seine Tiere von BIRKHAHN erhielt.
| Beschreibung | Einklappen |
| Größe Kleine Oophaga Art mit einer von 19-21 mm. |
Körper eher schlank und gestreckt. Kopf schmaler als Körper. Kopf etwa so breit wie lang. Schnauze stumpf abgerundet. Zähne fehlend. senkrecht elliptisch. Schallblase beim Ruf groß und deutlich sichtbar. Körper und Gliedmaßen grünlich, gelblich, türkis oder blau. Ein dunkles, braun bis schwarzbraunes Netzmuster auf dem Rücken ist mehr oder weniger stark ausgeprägt oder fehlt. Haut glatt. Finger 1 << Finger 2. Mit vergrößerten Haftscheiben. Haftscheibe an Finger 3 etwa 2-2,5 mal breiter als der Finger. Finger 1 besitzt nur schwach ausgeprägte Haftscheiben. Haftscheiben der Zehen kleiner, als die der Finger. Fingerformel: 3>4>2>1. Zehenformel: 4>3>5>2>1. Im Gegensatz zu Oophaga pumilio gibt es einen . Die Kehle der Männchen ist immer! dunkel gefärbt und lässt so eine sichere Geschlechtbestimmung zu, ein bei O. pumilio nicht immer sicheres Merkmal. (Alle Angaben nach JUNGFER, 1996)
Variation:
JUNGFER (1996) beschreibt vier Farbmorphen. Tiere von El Copé, der Terra typica, haben kaum Zeichnungselemente auf dem hellgrünen Körper. Tiere vom Río Concepción und dem Río Belén besitzen ein ausgeprägtes dunkelbraunes Netzmuster und eine grüne bis gelbgrüne Grundfarbe. Tiere aus El Valle de Antón besitzen eine sehr helle blassgrüne Färbung mit großen schwarzen Flecken und Bändern. Der Autor konnte noch eine weitere Variante im P.N. Santa Fé beobachten (OSTROWSKI, pers. Beobacht.). Dabei handelt es sich um die von BIRKHAHN 1996 nach Deutschland eingeführte Variante (CITES 2007). Die Tiere sind türkisgrün bis hellblau und zeigen darauf dunkelbraune Flecken und Streifen. MYERS & DALY (1976) und SILVERSTONE (1975) beschrieben als Dendrobates pumilio weitere Populationen aus den Provinzen Veraguas und der Comarca Ngöbe-Buglé (ehemals Bocas del Toro). Diese Populationen dürften, aufgrund ihrer von O. pumilio abweichenden Rufcharakteristika und ebenfalls O. vicentei zuzuordnen sein. Damit dürften es sich bei den Populationen vom Río Cahuita und dem Río Calovebora ebenfalls um O. vicentei handeln.
Weitere Varianten tauchten ab 2004 in Europa auf. Eine rote Morphe die dunkelbraun genetzt ist und eine dunkelbraune Morphe mit hellgrünen Flecken. Die Herkunft dieser Populationen und deren verwandtschaftliche Stellung ist noch nicht abschließend geklärt. Erste genetische Daten deuten für die rote Morphe auf eine Zugehörigkeit zum Taxon O. vicentei hin (s. o. Sytematik). Die gefleckte Morphe wurde fälschlicherweise als O. arborea bezeichnet. O. arborea stammt allerdings aus Westpanama und nicht aus Zentralpanama, wie die abgebildeten Tiere. O. arborea zeigt gegenüber den abgebildeten Tieren auch ein deutlich abweichendes Farbmuster mit gelben Flecken auf dunkelbraunen Grund und ist auch auf der Bauchseite dunkel gefärbt und gefleckt. Die abgebildeten Tiere zeigen dagegen eine helle ungezeichnete und eine zeichnung mit hellgrün bis mintfarbene Flecken auf dunkelbraunem Untergrund. Ihre Zeichnung ähnelt damit sehr stark der aus El Valle bekannten Morphe von O. vicentei.Das Vorkommen in Zantralpanama innerhalb des Verbreitungsgebietes von O. vicentei, die abweichende Färbung und die Rufcharakteristika lassen eher auf eine weitere Morphe von O. vicentei schließen. Bis zur abschließenden Klärung bezeichnen wir beide Varianten hier als O. cf. vicentei.
Weitere Varianten tauchten ab 2004 in Europa auf. Eine rote Morphe die dunkelbraun genetzt ist und eine dunkelbraune Morphe mit hellgrünen Flecken. Die Herkunft dieser Populationen und deren verwandtschaftliche Stellung ist noch nicht abschließend geklärt. Erste genetische Daten deuten für die rote Morphe auf eine Zugehörigkeit zum Taxon O. vicentei hin (s. o. Sytematik). Die gefleckte Morphe wurde fälschlicherweise als O. arborea bezeichnet. O. arborea stammt allerdings aus Westpanama und nicht aus Zentralpanama, wie die abgebildeten Tiere. O. arborea zeigt gegenüber den abgebildeten Tieren auch ein deutlich abweichendes Farbmuster mit gelben Flecken auf dunkelbraunen Grund und ist auch auf der Bauchseite dunkel gefärbt und gefleckt. Die abgebildeten Tiere zeigen dagegen eine helle ungezeichnete und eine zeichnung mit hellgrün bis mintfarbene Flecken auf dunkelbraunem Untergrund. Ihre Zeichnung ähnelt damit sehr stark der aus El Valle bekannten Morphe von O. vicentei.Das Vorkommen in Zantralpanama innerhalb des Verbreitungsgebietes von O. vicentei, die abweichende Färbung und die Rufcharakteristika lassen eher auf eine weitere Morphe von O. vicentei schließen. Bis zur abschließenden Klärung bezeichnen wir beide Varianten hier als O. cf. vicentei.
Tab.: Namen, Größenangaben und Farbmusterverteilung verschiedener Populationen von Oophaga vicentei
| Farbmorphe | Foto | Alternativ Namen | Verbreitung | Grundfarbe | Zeichnung | Größe [mm] |
| cf. vicentei |  | 22-23 | ||||
| El Copé |  | Prov. Coclé, Panama | grün, blau metallischer Ventralseite | ohne Zeichnung oder blasse braune Punkte | 18-20 | |
| El Valle |  | Prov. Coclé, Panama | mintfarben | schwarze Punkte | 18-20 | |
| Holotyp | ||||||
| Río Belén | Prov. Veraguas, Panama | gelb-grün | kleine braun bis schwarze Punkte und Bänder | 18-20 | ||
| Río Cahuita | ||||||
| Río Calovébora | ||||||
| Río Concepción |  | Prov. Veraguas, Panama | gelb-grün | braun bis schwarze Bänder | 18-20 | |
| Santa Fé |  | Prov. Veraguas, Panama | blau-türkis bis mintfarben | dunkelgrün bis braunes Wurmmuster auf dem Rücken | 18-20 |
Nicht bekannt, wahrscheinlich um die 5-10 Jahre.
Geschlechtsreife:
BRANDT (pers. Komm.) berichtet von einem Nachzuchtpaar, das nach ca. 12-14 Monaten die ersten Gelege absetzte. Das Männchen hatte etwa 2 Monate vorher zu rufen begonnen.
Hautgifte:
Bisher keine Angaben. Dürfte, wie bei den verwandten Arten des Gattung Oophaga, aus en und en bestehen.
| Gelege und Larven | Einklappen |
Merkmale Gelege:
Entwicklungszeiten:
Ernährung:
Legt nach JUNGFER ET AL. (1996) 2-6 Eier.
Die Larven schlüpfen nach 13-15 Tagen. Die ist nach 52-62 Tagen abgeschlossen. (JUNGFER ET AL., 1996)
Ernährung:
Wie auch O. pumilio . Die Larven nehmen ausser den arteigenen Nähreiern keine andere Nahrung auf.
| Verhalten (Ethologie) | Einklappen |
Allgemeines Verhalten:
Revierverhalten:
Balzverhalten:
Brutpflegeverhalten:
Lautäußerung (Vokalisation):
Ruf Oophaga vicentei, © 2005 T. Ostrowski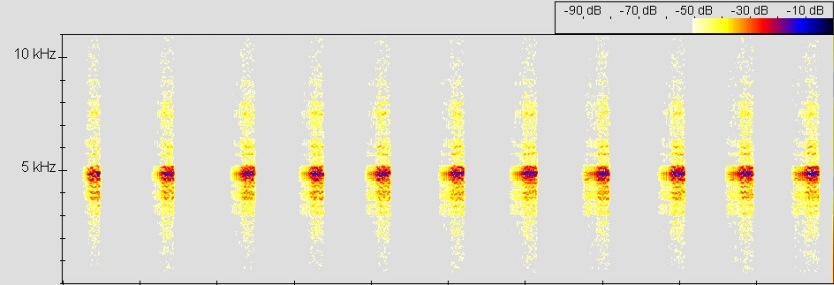
Sonagramm vergleichen:
Die Art ist streng und lebt hoch in den Bäumen in 5-15 m Höhe. Am Boden trifft man die Art nur sehr selten an. BIRKHAHN (pers. Komm.) fand auch einige wenige Tiere am Boden, während OSTROWSKI (pers. Beobacht.) und JUNGFER ET AL. (1996) keine Tiere am Boden nachweisen konnten. Das am tiefsten nachgewiesene Tier von JUNGFER befand sich 1,5 m über dem Boden. OSTROWSKI fand ein rufendes Männchen 2 m über dem Boden auf einem Palmblatt. Männchen rufen aus den Baumkronen in großer Zahl. Vom Verhalten ähnelt die Art stark O. pumilio.
Revierverhalten:
Über das Territorialverhalten liegen wenige Beobachtungen vor. Im Freiland leben die Tiere eher einzeln. OSTROWSKI konnte nie weitere Tiere in der unmittelbaren Nähe rufender Männchen antreffen. Im Terrarium sind die Männchen territorial und verhalten sich untereinander aggressiv (BRANDT, pers. Komm.). Damit ähneln die Art stark dem nah verwandten O. pumilio. Die ebenfalls stark arboricole O. arborea scheint wesentlich weniger aggressiv (MYERS & DALY, 1984).
Balzverhalten:
Auch im Balzverhalten zeigt sich, dass O. vicentei ein typischer Vertreter der Gattung Oophaga ist. Durch den Ruf werden paarungswillige (rezeptive) Weibchen angelockt. Bei Sichtkontakt wechselt das Männchen in einen anderen Ruftyp (s. Vokalisation). Wie bei anderen Arten von Oophaga, führt das Männchen, unter unregelmäßiger Abgabe des Werberufes, das Weibchen zum Eiablageort, meist die glatte Blattbasis von Bromelien. Dort wischt das Männchen einige Male mit den Hinterbeinen über das Blatt. Auch das Weibchen führt anschließend wischenden Bewegungen aus, während das Männchen in 1-2 cm Abstand verharrt. Das Weibchen dreht sich während der Wischbewegungen mit angedrücktem Bauch auf dem Blatt einige Male um sich selbst (Gegen- oder Uhrzeigersinn). Dabei wird das Blatt befeuchtet. Dieses Verhalten kann 30 min und mehr dauern. Das Männchen sitzt ruhig daneben und verhält sich meist sehr passiv. Schließlich klettert das Männchen zum Weibchen, bis beide Tiere entgegengesetzt Kloake an Kloake sitzen. In dieser Position wird vermutlich das Sperma abgegeben. Danach verlässt das Männchen den Ablageort. Oft ruft es nun wieder von benachbarten Pflanzenblättern. Das Weibchen legt nun die Eier ab, während es die Drehbewegungen fortsetzt (JUNGFER ET AL. , 1996 u. BRANDT, pers. Komm.). Dieses Verhalten mit getrennter Sperma- und Eiabgabe ist typisch für alle Vertreter der Unterfamilie Dendrobatinae.
Brutpflegeverhalten:
Nach JUNGFER ET AL. (1996) bewässert das Männchen während der 13-15 Tage dauernden Gelegephase mind. einmal am Tag das Gelege. Nach dem Schlupf nimmt das Weibchen die Larven auf den Rücken auf und transportiert die Larven einzeln in Wasser gefüllte ta. Das Weibchen besucht im Abstand von 2-8 Tagen jede Larve um abzulegen. Die Larve führt dazu schlängelnde Bewegungen aus sobald das Weibchen am Rand der Bromelienachsel auftaucht. Das Weibchen klettert dann rückwärts in die Achsel und legt 2-6 unbefruchteten Eier ab. Dabei schwimmt die Larve heftig um und gegen das Weibchen. Während das Weibchen Larven mit Nähreiern versorgt, setzt es keine weiteren befruchteten Gelege ab. Das Brutpflegeverhalten ähnelt damit sehr stark dem von O. pumilio (für genaue Angaben siehe dort).
Lautäußerung (Vokalisation):
Bei entsprechender Witterung rufen Männchen von O. vicentei den ganzen Tag über. Die meisten Männchen kann man im Biotop in den Morgenstunden bis 12.00 Uhr und in den Nachmittagsstunden von 15.00 bis 17.00 Uhr vernehmen. Bei sehr heißem Wetter lässt die Rufaktivität mittags nach. Bei starkem Regen oder Wind und auch bei sehr trockenem Wetter stellen die Tiere das Rufen ein.
Der Ruf von O. vicentei ist ein langsamer, insektenartiger und harscher (chirp-call). Wie alle Rufe von Arten der Unterfamilie Dendrobatinae besteht eine Rufsequenz aus einer Folge gepulster n ohne deutliche Frequenzmodulation. Man kann bei O. vicentei zwei Ruftypen unterscheiden. Man unterscheidet Ruftyp 1, einen (advertisement call) und Ruftyp 2, einen (close range mating call). Diese beiden Ruftypen sind typisch für alle Arten von Oophaga. (MYERS & DALY, 1984 u. JUNGFER, 1985). Ruftyp 1 besteht aus Noten, die zu Beginn und Ende einer oft unregelmäßig sind. Innerhalb des Rufes werden die Noten aber sehr regelmäßig abgegeben. Die Noten wiederum bestehen aus en mit ansteigender Amplitude zum Notenende hin (-> Gesamtes Sonagramm). Zu Beginn einer Note sind die Pulse recht unregelmäßig mit längeren , zum Ende aber ohne Intervall und sehr regelmäßig. Eine von OSTROWSKI vermessene Rufsequenz zeigte folgende Parameter: Der Frequenzbereich liegt zwischen 1,77 und 8,6 kHz. Die Dominanzfrequenz, also der höchste Energieanteil des Rufes, liegt bei 4,91 kHz (n = 44). Eine Note ist im Mittel 200 ms lang und die einer Rufsequenz liegt im Mittel bei 2,08 Noten/s (n = 68). Jede Note unterteilt sich in 28 – 31 teilweise nur schwach getrennte Pulse ( Ø = 29,4 Pulse; n = 10). Die liegt zwischen 140 – 158 Pulsen/s (Ø = 150,55 Pulse/s; n = 10). Zu Beginn einer Note sind die Pulse durch ein Interpulsintervall von 1,3 - 8,8 ms Dauer (Ø = 5,5 ms; n = 35) noch gut getrennt und haben eine Länge von 6,3 – 8,3 ms (Ø = 7,2 ms; n = 35). Am Ende der Note sind die Pulse kaum noch getrennt und wesentlich kürzer. Interpulsintevall: 0 - 1,1 ms (Ø = 0,05 ms; n = 65); Pulsdauer: 4 - 6,8 ms (Ø = 5,6 ms; n = 65). Ruftyp 2 unterscheidet sich nach JUNGFER (1996) von Ruftyp 1 durch eine etwas niedrigere Frequenz und eine geringere Rufrate. Die Notendauer beträgt 80 – 131 ms (Ø = 100,25 ms; n = 12) und die Dominazfrequenz liegt im Mittel bei 4,5 kHz. Durch die geringere Notendauer und geringere Rufrate erscheint der Werberuf von O. vicentei langsamer und kürzer als der Anzeigeruf.
Der Ruf von O. vicentei ist ein langsamer, insektenartiger und harscher (chirp-call). Wie alle Rufe von Arten der Unterfamilie Dendrobatinae besteht eine Rufsequenz aus einer Folge gepulster n ohne deutliche Frequenzmodulation. Man kann bei O. vicentei zwei Ruftypen unterscheiden. Man unterscheidet Ruftyp 1, einen (advertisement call) und Ruftyp 2, einen (close range mating call). Diese beiden Ruftypen sind typisch für alle Arten von Oophaga. (MYERS & DALY, 1984 u. JUNGFER, 1985). Ruftyp 1 besteht aus Noten, die zu Beginn und Ende einer oft unregelmäßig sind. Innerhalb des Rufes werden die Noten aber sehr regelmäßig abgegeben. Die Noten wiederum bestehen aus en mit ansteigender Amplitude zum Notenende hin (-> Gesamtes Sonagramm). Zu Beginn einer Note sind die Pulse recht unregelmäßig mit längeren , zum Ende aber ohne Intervall und sehr regelmäßig. Eine von OSTROWSKI vermessene Rufsequenz zeigte folgende Parameter: Der Frequenzbereich liegt zwischen 1,77 und 8,6 kHz. Die Dominanzfrequenz, also der höchste Energieanteil des Rufes, liegt bei 4,91 kHz (n = 44). Eine Note ist im Mittel 200 ms lang und die einer Rufsequenz liegt im Mittel bei 2,08 Noten/s (n = 68). Jede Note unterteilt sich in 28 – 31 teilweise nur schwach getrennte Pulse ( Ø = 29,4 Pulse; n = 10). Die liegt zwischen 140 – 158 Pulsen/s (Ø = 150,55 Pulse/s; n = 10). Zu Beginn einer Note sind die Pulse durch ein Interpulsintervall von 1,3 - 8,8 ms Dauer (Ø = 5,5 ms; n = 35) noch gut getrennt und haben eine Länge von 6,3 – 8,3 ms (Ø = 7,2 ms; n = 35). Am Ende der Note sind die Pulse kaum noch getrennt und wesentlich kürzer. Interpulsintevall: 0 - 1,1 ms (Ø = 0,05 ms; n = 65); Pulsdauer: 4 - 6,8 ms (Ø = 5,6 ms; n = 65). Ruftyp 2 unterscheidet sich nach JUNGFER (1996) von Ruftyp 1 durch eine etwas niedrigere Frequenz und eine geringere Rufrate. Die Notendauer beträgt 80 – 131 ms (Ø = 100,25 ms; n = 12) und die Dominazfrequenz liegt im Mittel bei 4,5 kHz. Durch die geringere Notendauer und geringere Rufrate erscheint der Werberuf von O. vicentei langsamer und kürzer als der Anzeigeruf.
Ruf Oophaga vicentei, © 2005 T. Ostrowski
Sonagramm vergleichen:
| Lebensraum | Einklappen |
Typenfundort der Erstbeschreibung
Daten von OpenStreetMap - Veröffentlicht unter CC-BY-SA 2.0
Biotop:
Klima:
"Aus dem Fussweg von El Copé nach Río Blanco del Norte, Passhöhe der Kontinentlscheide etwa 1 km östlich des Cerro Blanco, etwa 8° 40´ N, 80° 36´ W, 912 m NN, Porvincia de Coclé, Panamá".
JUNGFER ET AL. (1996)
JUNGFER ET AL. (1996)
Verbreitung: |  Abb.: Verbreitungsgebiet Oophaga vicentei © 2008 () |
Biotop:
Wie auch O. arborea besiedelt O. vicentei die Passhöhen der Cordilleren und auch die karibischen Tieflandwälder. Es handelt sich um feuchte, heiße Tieflandregenwälder oder feuchte, warme e Bergregenwälder. Die Tiere leben im Stamm- und Kronenbereich von hohen Bäumen in 4 – 15 m Höhe. Die Bäume in höheren Lagen stehen dabei nicht mehr ganz so dicht und sind nicht so hoch, wie die Bäume der Tieflandwälder. Die Äste bewohnter Bäume sind aber immer dicht mit besetzt. Im Bereich des N.P. Santa Fé waren Bromelien der Gattung Guzmania ssp. typisch für die Epiphytenflora der Bäume (Abb.:). Tiere fanden sich aber nicht nur in der Nähe von Bromelien sondern auch auf Palmblättern, Lianen und bemoosten Ästen (OSTROWSKI, pers. Beobacht.). JUNGFER ET AL. (1996) berichten von einem Steilhang im P.N. Omar Torrijos Herrera (El Copé) der mit Bromelien bewachsen war und dort das Biotop von O. vicentei darstellte. Im Bereich von El Copé und Santa Fé leben Oophga vicentei mit Dendrobates auratus, Ranitomeya minuta, Silverstoneia nubicola, S. cf. pratti, S. flotator und Phyllobates lugubris. Durch die streng arboricole Lebensweise von O. vicentei begegnet er den anderen Arten normalerweise nicht, da diese in der Laubstreu oder an Fließgewässern völlig andere ökologische Nischen besetzen. Im Bereich des Río Concepcion soll es nach Aussage von REICHELT (pers. Komm.) auch ein sympatrisches Vorkommen mit dem nah verwandten O. pumilio geben.

Abb.: Karibische und pazifischen Kammregionen der Cordillera Central sind das Hauptverbreitungsgebiet von O. vicentei.
© 2005 Thomas Ostrowski
Abb.: Karibische und pazifischen Kammregionen der Cordillera Central sind das Hauptverbreitungsgebiet von O. vicentei.
© 2005 Thomas Ostrowski
 Abb.: Die karibischen Hänge der Cordilera Central sind schwer zugänglich und in der Regenzeit nur zu Fuß erreichbar. © 2005 Thomas Ostrowski |
 Abb.: Viehweide mit alten Überständern bei Alto Piedra. Wohnbäume von O. vicentei. © 2005 Thomas Ostrowski |
 Abb.: Unberührter Bergregenwald im Nationalpark Santa Fé. © 2005 Thomas Ostrowski |
 Abb.: Guzmania sp., typische Brutbromelie von O. vicentei. © 2005 Thomas Ostrowski . |
 Abb.: Biotop Oophaga vicentei © 2008 Thomas Ostrowski |
 Abb.: Biotop Oophaga vicentei © 2008 Thomas Ostrowski |
 Abb.: Biotop Oophaga vicentei © 2008 Thomas Ostrowski |
 Abb.: Biotop Oophaga vicentei © 2008 Thomas Ostrowski |
Klima:
Oophaga vicentei ist eine Tieflandart die bis zu den Passhöhen der Cordilleren aufsteigt. Im Tiefland herrscht feuchtheißes, relativ konstantes Klima mit Temperaturen von 25-27 °C mit einer mittleren Jahresschwankung von nur 1-2 °C. Der Tieflandregenwald zeichnet sich auch durch rel. geringe Tagesschwankungen von 4-5 °C aus. Mit zunehmender Höhe sinkt die mittlere Jahrestemperatur nach WALTER & BRECKLE (1999) alle 200 m um 1,2 °C und erreicht auf den Passhöhen der Cordillera Central somit nur noch etwa 22°C am Tag. Die Tagesschwankungen fallen hier mit durchschnittlich 5-8 °C und in Extremfällen 10 °C schon recht hoch aus. Nachts kann es empfindlich kalt werden und die Temperatur bis auf 12 °C fallen. Wir konnten O. vicentei jedoch nur in klimatisch begünstigten Biotopen nachweisen, in denen durch Berghänge geschützt auch in 600 m Höhe noch Tieflandtemperaturen von 24-26 °C herrschten (Alto Piedra). JUNGFER ET AL.(1996) berichtet ebenfalls von „...nicht zu niedrigen Temperaturen des 912 m hohen Passes..“ der Terra typica. Sicher ist jedoch ein Wipfelbiotop stärkeren Temperaturschwankungen ausgesetzt als ein Biotop in der Laubstreu am Waldboden. Während in der Laubschicht auch nachts die Temperatur nur wenig sinkt, und unter der Laubdecke fast ganzjährig die mittlere Jahresdurchschnittstemperatur gemessen werden kann, treten im Kronenbereich (unabhängig von der Höhe) durch Starkregen, Winde und Nachtkühle starke Temperaturunterschiede in rel. kurzer Zeit auf. Solche starken Temperaturschwankungen scheinen im Terrarium auch das Ruf- und damit Balzverhalten auszulösen (BRANDT, pers. Komm.). Tiere die an klimatisch begünstigten Passhöhen bis auf die Pazifikseite vordringen, wie bei Santa Fé, El Copé und in El Valle, sind in den Monaten Januar bis April einer Trockenperiode mit wenig Niederschlag ausgesetzt(s. Abb.). Die karibischen Hänge der Cordillera erhalten ganzjährig viel Niederschlag.
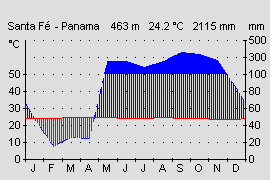 Abb.: Klimadiagramm der Pazifikseite Panamas. | 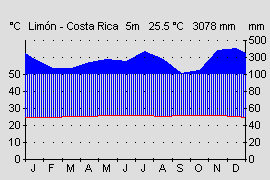 Abb.: Typisches, dauerfeucht und heißes Klima der Atlantikseite Mittelamerikas am Beispiel Limón, Costa Rica. |
| Haltung im Terrarium | Einklappen |
Terrarium/Einrichtung:
Temperaturen:
Luftfeuchte:
Ernährung:
Besatz:
Tipps zur Zucht:
Varianten in Terrarienhaltung:
Regenwald-Terrarium ab 50cm x 50 cm x 65cm. Das Becken sollte recht hoch sein. Kletteräste in großer Anzahl, da die Tiere viel klettern. Eine größere Bromelie als Reviermittelpunk sollte vorhanden sein.
Temperaturen:
Je nach Fundort nicht zu warm halten! 24-26°C für Tieflandpopulationen mit geringer Nachtabsenkung von 2-4 °C. Tiere aus dem Hochland bei 22-24 °C, nachts um 4-6 °C absenken. Im Freiland können die Temperaturen auch im Hochland am Tage 26 –28 °C erreichen, in 15 m Höhe herrscht aber eine gute Luftumwälzung, so dass die Tiere selten solchen Temperaturen ausgesetzt sind. Nachts sinken die Temperaturen auf 800 m manchmal bis unter 15 °C! Jahrestemperaturschwankung minimal (1-2 °C).
Luftfeuchte:
Nicht zu feucht halten! 70-85%, fällt zur Mittagszeit bis auf 70%, morgens und abends 100% (Nebel). Da die Tiere entlang der Wasserscheide leben und hier immer feuchte Luft von der Atlantikseite auftrifft gibt es keine ausgeprägte Trockenzeit. Tiere von etwas tieferen Lagen der Pazifikseite sind jedoch von Dezember bis April einer Trockenperiode ausgesetzt, wobei in den Achseln der großen Bromelien immer etwas Wasser zu finden ist.
Die starken Temperatur und Feuchtigkeitsschwankungen sind für eine optimale Haltung Vorraussetzung!!! BRANDT (pers. Komm.) berichtet von Temperaturstürzen als Auslöser für das Balzverhalten
Ernährung:
Übliche kleine Futtertiere , Mikro-Heimchen, kleinste Wachsmaden, Wiesenplankton und . Jungtiere sind sehr klein, bewältigen aber laut JUNGFER ET AL. (1996) im Gegensatz zu O. pumilio aber sofort Drosophila
Besatz:
Die Art wird selten gehalten. Doch wie in der Natur sollte eine optimale Gruppe aus 1,1 bestehen. Jungtiere werden lange in der Wohnbromelie geduldet.
Tipps zur Zucht:
Die Eiablage erfolgt in den Blattachsel der Wohnbromelie.
Legt 4-6 Eier
Entwicklungsdauer Eier: 12 -16 Tage
Die geschlüpften Quappen werden getrennt in den Achseln der Wohnbromelie verteilt.
Sie sind untereinander sehr aggressiv.
Entwicklungsdauer der Quappen: 80-90 Tage, Wassertemperatur bei 24°C, Nachtabsenkung empfohlen.
Es handelt sich bei den Quappen wie bei allen Fröschen der Histrionicus- Gruppe um Eierfresser. Das Weibchen legt auch hier spezielle Futtereier. Die Aufzucht ohne diese ist kaum möglich. Die Jungen verbleiben am besten bei den Eltern bis sie von den Eltern vertrieben werden. Dieses erfolgt oft erst nach mehreren Monaten. Oft verlassen die Jungen aber schon eher die Wohnbromelie um eine eigene Wohnstatt zu finden.
(Text copyright 2003 Thomas Ostrowski)
Legt 4-6 Eier
Entwicklungsdauer Eier: 12 -16 Tage
Die geschlüpften Quappen werden getrennt in den Achseln der Wohnbromelie verteilt.
Sie sind untereinander sehr aggressiv.
Entwicklungsdauer der Quappen: 80-90 Tage, Wassertemperatur bei 24°C, Nachtabsenkung empfohlen.
Es handelt sich bei den Quappen wie bei allen Fröschen der Histrionicus- Gruppe um Eierfresser. Das Weibchen legt auch hier spezielle Futtereier. Die Aufzucht ohne diese ist kaum möglich. Die Jungen verbleiben am besten bei den Eltern bis sie von den Eltern vertrieben werden. Dieses erfolgt oft erst nach mehreren Monaten. Oft verlassen die Jungen aber schon eher die Wohnbromelie um eine eigene Wohnstatt zu finden.
(Text copyright 2003 Thomas Ostrowski)
Varianten in Terrarienhaltung:
| cf. vicentei |  |
| El Copé |  |
| El Valle |  |
| Holotyp |  |
| Río Belén |  |
| Río Cahuita |  |
| Río Calovébora |  |
| Río Concepción |  |
| Santa Fé |  |
Daten errechnet aus von Nutzern gemeldeten Fröschen (Home->Meine Frösche) und den aktuellen Inseraten.
stand 18.03.2024 01:05 Uhr
stand 18.03.2024 01:05 Uhr
| Autoren dieser Seite: | Einklappen |
| Referenzen | Einklappen |
CITES (2005): Convention on International Trade of Endangered Species of Wild Fauna and Flora.
XXOnlineversion: 11.08.05. Electronic Database accessible at http://www.cites.org/index.html
FROST, D.R. (2007): Amphibian Species of the World: an Online Reference. Version 5.0 (1.02.2007).
XXElectronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.php
XX Am.Mus.Nat.Hist. New York, USA.
IUCN, Conservation International, and NatureServe. (2009): Global Amphibian Assessment.
XXwww.globalamphibians.org. Downloaded on 15 October 2004.
JUNGFER, K.H. (1985): Beitrag zur Kenntnis von Dendrobates speciosus O. SCHMIDT, 1857.
XX- Salamandra, Bonn, 21 (4): 263-280.
JUNGFER, K.H. ,WEYGOLD, P. & N. JURASKE (1996): Dendrobates vicentei, ein neuer Pfeilgiftfrosch
XXaus Zentral Panama. - herpetofauna, Weinstadt, 18(103): 17-26
MYERS, C.W. DALY, J.W. & MARTINEZ, V. (1984): An arboreal poison frog (Dendrobates) from
XXwestern Panama.- Am.Mus.Nov. 2783: 1-20.
XXOnlineversion: 11.08.05. Electronic Database accessible at http://www.cites.org/index.html
FROST, D.R. (2007): Amphibian Species of the World: an Online Reference. Version 5.0 (1.02.2007).
XXElectronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.php
XX Am.Mus.Nat.Hist. New York, USA.
IUCN, Conservation International, and NatureServe. (2009): Global Amphibian Assessment.
XXwww.globalamphibians.org. Downloaded on 15 October 2004.
JUNGFER, K.H. (1985): Beitrag zur Kenntnis von Dendrobates speciosus O. SCHMIDT, 1857.
XX- Salamandra, Bonn, 21 (4): 263-280.
JUNGFER, K.H. ,WEYGOLD, P. & N. JURASKE (1996): Dendrobates vicentei, ein neuer Pfeilgiftfrosch
XXaus Zentral Panama. - herpetofauna, Weinstadt, 18(103): 17-26
MYERS, C.W. DALY, J.W. & MARTINEZ, V. (1984): An arboreal poison frog (Dendrobates) from
XXwestern Panama.- Am.Mus.Nov. 2783: 1-20.
| Zitierung: | Einklappen |
OSTROWSKI, T. & T. MAHN (2024): Artbeschreibung Oophaga vicentei.
- Dendrobase.de - Eine Online-Datenbank der Familie Dendrobatidae (Anura).
https://www.dendrobase.de/html/D_oophaga_vicentei.html; Stand: März 2024
- Dendrobase.de - Eine Online-Datenbank der Familie Dendrobatidae (Anura).
https://www.dendrobase.de/html/D_oophaga_vicentei.html; Stand: März 2024
| Link für Website Betreiber: | Einklappen |
Webmaster dürfen Links von Ihren Seiten zur DendroBase erstellen, wenn sie folgende Kriterien erfüllen:
- Die Website enthält weder illegale, rassistische oder sexistische Inhalte
- Das Thema der Website bezieht sich auf Frösche
- Der Link genau wie folgend angegeben verwendet wird:
<a href="https://www.dendrobase.de/html/D_oophaga_vicentei.html" target="_new">Oophaga vicentei (DendroBase.de)</a>
- Die Website enthält weder illegale, rassistische oder sexistische Inhalte
- Das Thema der Website bezieht sich auf Frösche
- Der Link genau wie folgend angegeben verwendet wird:
<a href="https://www.dendrobase.de/html/D_oophaga_vicentei.html" target="_new">Oophaga vicentei (DendroBase.de)</a>
| Fotos | Einklappen |

Oophaga vicentei
"El Copé"
© 2003 Holger Birkhahn

Oophaga vicentei
"Santa Fé"
© 2004 ()

Oophaga vicentei
"Santa Fé"
© 2003 Holger Birkhahn

Oophaga vicentei
"Santa Fé"
© 2004 ()

Oophaga vicentei
"Río Concepción"
© 2005 ()

Oophaga vicentei
"Río Concepción"
© 2005 ()

Oophaga vicentei
© 2005 Ian Rogers

Oophaga vicentei
© 2005 Ian Rogers

Oophaga vicentei
"El Valle"
© 2008 Max Bjørneskov

Oophaga vicentei
"El Valle"
© 2008 Max Bjørneskov

Oophaga cf. vicentei
© 2005 Ian Rogers

Oophaga cf. vicentei
© 2005 Ian Rogers

Oophaga cf. vicentei
© 2005 ()

Oophaga cf. vicentei
© 2008 ()

Oophaga cf. vicentei
© 2008 ()

Oophaga cf. vicentei
© 2008 ()

Oophaga cf. vicentei
© 2008 ()

Oophaga cf. vicentei
© 2008 ()

Oophaga cf. vicentei
© 2008 ()

Oophaga cf. vicentei
© 2008 ()
DendroBase.de Version 5 | Impressum | Datenschutzerklärung