| Login | Einklappen |
| Fotos von Nutzern | Einklappen |

R. lamasi mit Quappe
© 2012 ()

Ranitomeya lamasi mit Quappe
© 2013 ()

Ranitomeya lamasi "Quimpichan"
© 2011 ()

R.lamasi, Gruppenfoto samt noch nicht ausgeschlüpftem Nachwuchs
© 2012 ()

R. lamasi Weibchen
© 2012 ()

R.lamasi Männchen
© 2012 ()

R. lamasi mit Quappe
© 2012 ()

R.lamasi mit Quappe
© 2013 ()

lamasi-Quappe kurz vor Landgang
© 0000 ()
| Nomenklatur | Einklappen |
Etymologie:
Synonyme:
Ranitomeya lamasi (GRANT, FROST, CALDWELL, GAGLIARDO, HADDAD, KOK, MEANS, NOONAN, SCHARGEL & WHEELER, 2006)
Dendrobates lamasi (MORALES, 1992)
sensu FROST, 2006.
englischer Name: Pasco Poison Frog
deutscher Name: Pasco Baumsteiger
| Systematik | Einklappen |
Amphibia->Anura->Dendrobatoidea->Dendrobatidae->Dendrobatinae->Ranitomeya->Ranitomeya lamasi (MORALES, 1992)
Gruppe:
Anmerkung zur Systematik:
Die in Terrarien verbreitete Population von der Divisoria der Cordillera Azul (Montanform nach SCHULTE, 1999) weicht sowohl in der Größe als auch vom Zeichnungsmuster etwas vom ab. Der von Tingo Maria (MORALES, 1992) ähnelt im Zeichnungsmuster sehr den Tieren einer Population, die aus der Umgebung der Forschungsstation Panguana am Río Llullapichis zunächst als D. quinquevittatus (MEEDE, 1980) und dann als D. sp. "Panguana" bekannt geworden ist. Übereinstimmungen in der und Vergleiche der Rufparameter bzw. Sonagramme von R. lamasi (MORALES, 1992b u.) und R. sp. "Panguana" (MEEDE, 1992 und OSTROWSKI & MAHN, 2005) weisen stark auf die Zugehörigkeit zu einem einzigen hin. Als morphologische Gemeinsamkeit zeigen sowohl die Hochlandform, als auch die Tiere von Tingo Maria und die Panguana Population typische Signalflecken in den Arm und Beinbeugen. Weitere Übereinstimmungen zeigen sich in der Larvenmorphologie. Das bei SCHULTE (1999) beschriebene metallisch grüne oder gelbe Querband auf der Schnauzenspitze von R. lamasi wird auch von Larven der "Panguana"- und der Hochlandpopulation gezeigt. Es dürfte sich also bei diesen Populationen ebenfalls um R. lamasi handeln. Weitere Fundorte von morphologisch sehr ähnlichen Populationen, sind bekannt. So konnten 2004 bei Contamana am Río Ucayali Tiere nachgewiesen werden, die ebenfalls in Adult- und Larvenmorphologie nahezu identisch mit R. lamasi sind (BIRKHAHN, pers. Komm.). Im Jahr 2005 wurden auch am Río Pachitea weitere Varianten nachgewiesen (NIESZPOREK, pers. Komm.), die in Ruf und Morphologie den übrigen Populationen gleichen und wahrscheinlich ebenfalls R. lamasi zuzuordnen sind. Bis weitere Rufanalysen von allen Populationen und genetische Vergleichsdaten diese Vermutung untermauern, bezeichnen wir die Tiere der Río Pachitea Drainage hier zunächst als Ranitomeya cf. lamasi.
Achtung!: Die Panguana Population wird mitunter als R. imitator „Panguana“ bezeichnet. Verbreitung, ethologische und morphologische Daten und die Rufanalyse (s. Vergleichsfunktion unter Ruf) dürften die Zugehörigkeit zum Taxon R. imitator jedoch ziemlich sicher ausschließen.
| Bedrohungsstatus | Einklappen |
Auf der Roten Liste ist die Art als nicht gefärdet eingestuft. Trotz des kleinen Verbreitungsgebietes von weniger als 20 000 qkm gilt der Lebensraum als intakt und die Populationen gelten als stabil (IUCN, 2004). Laut CITES (2005) ist ein Export aus Peru illegal und kommerzieller Handel nicht bekannt.
| Beschreibung | Einklappen |
| Größe Kleine Pfeilgiftfrosch-Art mit einer von 17-22 mm. Die Population vom Typenfundort und die Panguana Population rangieren eher am unteren Ende der Skala, während die Montanform und einige Tieflandvarianten (Contamana) eher um die 20 mm groß sind. |  © 2008 Thorsten Mahn |

Abb.1: Ranitomeya lamasi. Dorsal- und Ventralansicht des Paratypus aus Tingo Maria, MHNJP(=MHNSM) 1462. Orginalzeichnung aus MORALES, 1992.
Die Haut ist glatt mit schwarzer Grundfarbe. Typisch für R. lamasi sind nach MORALES (1999) die drei schmalen Längsstreifen auf dem Rücken in gelb, gold metallisch, orange, rot oder hellgrün. Die Streifen können bei einigen Populationen verbreitert und im unteren Rückenbereich miteinander verbunden sein (z.B. "Divisoria"). Der des Rückens bildet vor den Augen auf der Schnauzenspitze ein T oder Y. Der Schwarze Nasenbalken auf der Schnauzenspitze ist manchmal nur auf einen runden Fleck reduziert. Entlang der Flanke zieht sich ein weiterer Streifen in gleicher Farbe wie die . Dieser zieht unterhalb des Auges um die Schnauze herum und kann auf der Flanke ebenfalls mit dem verbunden sein (siehe Zeichnungen). Die Beine zeigen ein Flecken-Netzwerk in grau weiß oder blau auf schwarzem Grund. Auf der Oberseite am Ansatz des Oberarmes und des Oberschenkels befindet sich ein weiß bis hellgelber Signalfleck. Die Tiere zeigen ein typisches Bauchmuster. Dabei sind Kehle und Bauch kontrastierend zur Färbung der unteren Flanken der Beine in der Farbe der - und Dorsalstreifen gefärbt. Kehle und Bauchfleck sind wiederum durch ein Band getrennt welches in Beinfarbe gezeichnet ist (vgl. Zeichnung und Fotos). Auch die nah verwandten Arten R. biolat, R. flavovittata und R. vanzolinii zeigen diese unverwechselbare Bauchzeichnung (Vanzolinii-Brustgürtel nach SCHULTE, 1999). Maxillarzähne (Oberkiefer) fehlend. Augen groß. kleiner als Augendurchmesser, zu 3/4 sichtbar. Finger mit verbreiterten Haftscheiben. Ohne Schwimmhäute oder Hautsäume an Finger und Zehen. : 1<2<4<3. : 1<2<5<3<4, Zeh 1 reduziert.
 Abb.2: Ranitomeya lamasi. Paratyp MHNJP(=MHNSM) 1462; Seitenansicht des Kopfes. Orginalzeichnung aus MORALES, 1992. |
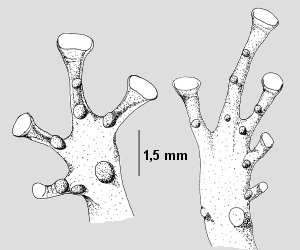 Abb.3: Ranitomeya lamasi. Paratyp MHNJP(=MHNSM) 1462; Ventrale Ansicht der linken Vorderhand (links) und des rechten Hinterfußes (rechts). Orginalzeichnung aus MORALES, 1992. |
 Abb.: Dorsal- und Ventralansicht Ranitomeya cf. lamasi "Panguana" copyright 2006 () | Größe vergleichen: Dorsal- und Ventralbild vergleichen: Hand- und Fußabbildung vergleichen: |
Variation:
| Ranitomeya lamasi "Tingo Maria" |  |  |
| Ranitomeya lamasi "Divisoria" | |  |
| Ranitomeya cf. lamasi "Panguana" | |  |
| Ranitomeya cf. lamasi "Contamana" |  |  |
| Ranitomeya cf. lamasi |  |  |
| Ranitomeya cf. lamasi |  |  |
Alter:
Geschlechtsreife:
Hautgifte:
| Gelege und Larven | Einklappen |
Merkmale Larven:

Abb.5: Larve von R. lamasi kurz vor der Metamorphose.
© 2006 Wojtek Nieszporek
 Abb.: Larve Ranitomeya cf. lamasi © 2005 () |
Entwicklungszeiten:
Ernährung:
| Verhalten (Ethologie) | Einklappen |
Revierverhalten:
Balzverhalten:
Brutpflegeverhalten:
Lautäußerung (Vokalisation):
Ruf Ranitomeya cf. lamasi "Panguana", © 2005 T. Mahn
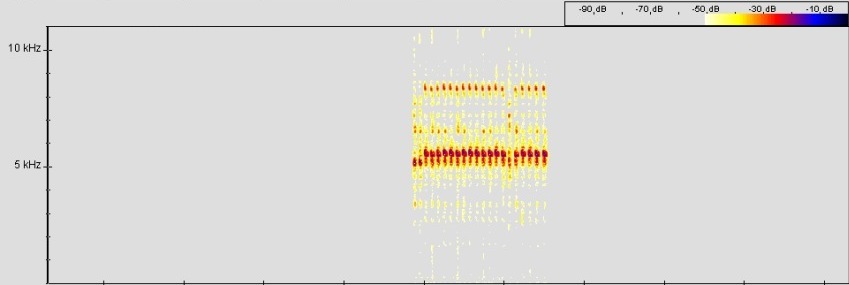
Abb.: Spectrogram, FFT size 512, Hanning window
Sonagramm vergleichen:
| Lebensraum | Einklappen |
Verbreitung: | 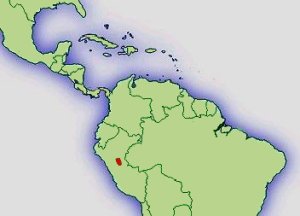 Abb.: Verbreitungsgebiet Ranitomeya lamasi © 2008 () |
Biotop:
 Abb.: Entlang der Pfade im Biotop bilden sich dichte Bestände von Stauden. Die Blattscheiden verschiedener Arten werden von R. lamasi als Brutpflanzen genutzt. © 2006 W. Nieszporek |
 Abb.: R. cf. lamasi an einer Xanthosoma sp. im Biotop. Departamento Huánuco, Peru © 2006 W. Nieszporek |
 Abb.: Als Brutpflanzen werden von Ranitomeya cf. lamasi auch die wassergefüllten Blattscheiden von Heliconia sp. genutzt. Departamento Huánuco, Peru. © 2008 Thomas Ostrowski |
 Abb.: Ranitomeya cf. lamasi bewohnt im Primärwald oft Lianen direkt oberhalb der Krautschicht. Departamento Huánuco, Peru. © 2008 Thomas Ostrowski |
 Abb.: Biotop von Ranitomeya cf. lamasi entlang einer kleinen Quebrada am oberen Río Pachitea. Departamento Huánuco, Peru. © 2008 Thomas Ostrowski |
 Abb.: Am Waldboden im Biotop findet sich Ranitomeya cf. lamasi nur selten. Departamento Huánaco, Peru. © 2008 Thomas Ostrowski |
Klima:
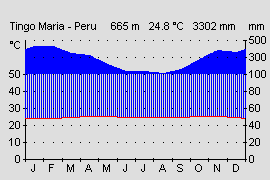 Abb.: Klimadiagramm der submontanen Stufe am Beispiel Tingo Maria, Peru. | 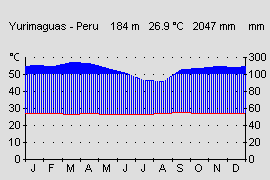 Abb.: Klimadiagramm der prämontanen Stufe des Tieflandregenwaldes am Beispiel Yurimaguas, Peru. |
| Haltung im Terrarium | Einklappen |
Temperaturen:
Luftfeuchte:
Ernährung:
Besatz:
Tipps zur Zucht:

© 2008 Thomas Ostrowski
Varianten in Terrarienhaltung:
| Contamana |  |
| Divesoria |  |
| Fundort |  |
| Holotyp |  |
| Panguana |  |
| Puerto Inca |  |
| Quimpichari |  |
| Yuyapichis |  |
stand 02.04.2024 08:30 Uhr
| Autoren dieser Seite: | Einklappen |
| Referenzen | Einklappen |
XX revision of the quinquevittatus Group of Dendrobates. - Am.Mus.Nov. 2988: 1-21. (PDF)
CHRISTMANN, S.P. (2004): Dendrobatidae. Baumsteigerfrösche. Eine Phantastische Reise durch
XXEcuador-Peru-Kolumbien. Bd. II. - Liebl Druck München.
CITES (2005): Convention on International Trade of Endangered Species of Wild Fauna and Flora.
XXOnlineversion: 11.08.05. Electronic Database accessible at http://www.cites.org/index.html
FROST, D.R. (2007): Amphibian Species of the World: an Online Reference. Version 5.0 (1.02.2007).
XXElectronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.php
XX Am.Mus.Nat.Hist. New York, USA.
GOSNER, K.L. (1960): A Simplified Table for Staging Anuran Embryos and Larvae.
XX-Herpetologica, 16: 183-190
GRANT, T . , FROST, D. R. , CALDWELL, J. P. , GAGLIARDO, R. , HADDAD, C. F. B. , KOK, P. J. R. ,
XXMEANS, D.B. , NOONAN, B. P. , SCHARGEL, W. E. & W.C. WHEELER (2006): Phylogenetic
XXsystematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae).
XX - Bulletin of the Am.Mus.Nat.Hist. 299. (PDF)
IUCN, Conservation International, and NatureServe. (2009): Global Amphibian Assessment.
XXwww.globalamphibians.org. Downloaded on 15 October 2004.
MEEDE, U. (1980): Beobachtungen an Dendrobates quinquevittatus und Phyllobates fermoralis.
XX- Salamandra, Frankfurt/M., 16(1): 38-51.
MORALES, V. R. (1992): Dos Especies Nuevas de Dendrobates (Anura: Dendrobatidae) para Peru.
XX- Caribbean Journal of Science. 28 (3-4): 191-199.
MORALES, V. R. (1992): Estudio de la vocalizacion de algunas ranas dardo-venenoso (Dendrobatidae,
XXDendrobates) en el Peru. - Acta Zool. Lilloana 41.
SCHULTE, R. (1999): Pfeilgiftfrösche - Artenteil Peru. - Nikola Verlag, Stuttgart.
SILVERSTONE, P.A. (1975): A Revision of the Poison-Arrow-Frogs of the Genus Dendrobates Wagler.
XX- NatHist.Mus. Los Angeles County, Sci.Bull. 21: 1-55. (PDF)
WALTER, H. & S. BRECKLE (1999): Vegetationen und Klimazonen. 7. Auflage. - Ulmer, Stuttgart.
| Zitierung: | Einklappen |
- Dendrobase.de - Eine Online-Datenbank der Familie Dendrobatidae (Anura).
https://www.dendrobase.de/html/D_ranitomeya_lamasi.html; Stand: April 2024
| Link für Website Betreiber: | Einklappen |
- Die Website enthält weder illegale, rassistische oder sexistische Inhalte
- Das Thema der Website bezieht sich auf Frösche
- Der Link genau wie folgend angegeben verwendet wird:
<a href="https://www.dendrobase.de/html/D_ranitomeya_lamasi.html" target="_new">Ranitomeya lamasi (DendroBase.de)</a>
| Fotos | Einklappen |

Ranitomeya cf. lamasi
"Divisoria"
© 2002 ()

Ranitomeya cf. lamasi
"Puerto Inca"
© 2004 Andreas Toebe

Ranitomeya cf. lamasi
"Puerto Inca"
© 2005 ()

Ranitomeya cf. lamasi "Puerto Inca"
© 2005 ()

Ranitomeya lamasi
"Yuyapichis"
© 2008 ()

Ranitomeya lamasi
"Yuyapichis"
© 2008 ()

Ranitomeya lamasi
"Quimpichari"
© 2008 ()

Ranitomeya lamasi
"Quimpichari"
© 2008 ()

Ranitomeya lamasi
"Quimpichari"
© 2008 ()

Ranitomeya cf. lamasi
© 2005 Wojtek Nieszporek

Ranitomeya cf. lamasi
© 2005 Wojtek Nieszporek

Ranitomeya cf. lamasi
Gleiches Zeichnungsmuster wie der Paratyp von Tingo Maria.
© 2006 Wojtek Nieszporek

Ranitomeya lamasi
"Contamana"
© 2009 Evan Twomey

Ranitomeya lamasi
"Contamana"
© 2009 Jason Brown