| Login | Einklappen |
Startseite
Artbeschreibungen
Allobates
Anomaloglossus
Rheobates
Aromobates
Mannophryne
Ameerega
Colostethus
Epipedobates
Silverstoneia
Andinobates
Adelphobates
Dendrobates
Excidobates
Minyobates
Oophaga
Phyllobates
Ranitomeya
Hyloxalus
Foren
Inserate
Reportagen
Service & Links
Einstellungen
| Fotos von Nutzern | Einklappen |

Ranitomeya ()uakarii Landgänger
© 0000 ()

Ranitomeya ()uakarii
© 0000 ()

Ran. ()uakarii
© 2013 ()
| Nomenklatur | Einklappen |
wissenschaftl. Name: Ranitomeya uakarii BROWN, SCHULTE & SUMMERS 2006
Etymologie:
Synonyme:
Dendrobates uakarii (BROWN, SCHULTE & SUMMERS 2006)
englischer Name: Tahuayo Poison Frog
deutscher Name: Tahuayo Baumsteiger
Etymologie:
Das uakarii bezieht sich auf eine im Biotop vorkommende rotköpfige Affenart. Den Kahlkopf-Uakari (Cacajao calvus). Gemeinsamkeiten in der Biogeografie und die kräftig rote Hautpigmentierung des Kopfes, die beiden
Arten gemeinsam ist, führte zur Namensgebung durch BROWN ET AL. (2006).
Synonyme:
Dendrobates uakarii (BROWN, SCHULTE & SUMMERS 2006)
englischer Name: Tahuayo Poison Frog
deutscher Name: Tahuayo Baumsteiger
| Systematik | Einklappen |
Klassifizierung:
Amphibia->Anura->Dendrobatoidea->Dendrobatidae->Dendrobatinae->Ranitomeya->Ranitomeya uakarii BROWN, SCHULTE & SUMMERS 2006
Anmerkung zur Systematik:
Amphibia->Anura->Dendrobatoidea->Dendrobatidae->Dendrobatinae->Ranitomeya->Ranitomeya uakarii BROWN, SCHULTE & SUMMERS 2006
Anmerkung zur Systematik:
Lange Zeit unter der Bezeichnung Dendrobates ventrimaculatus "Durchgestreift" in Haltung. Dann fälschlicherweise als Dendrobates duellmani bezeichnet. Anfang 2006 wurden die Tiere zunächst als eigene Art beschrieben (BROWN ET AL., 2006) und dann von GRANT ET AL. (2006) aus der Gattung Dendrobates in die Gattung Ranitomeya überstellt. Von Ranitomeya duellmani unterscheidet sich Ranitomeya uakarii in einigen morphologischen und ethologischen Merkmalen.
Ob einige ähnlich wie R. uakarii gefärbte Populationen am Río Pachitea und Río Napo, hier als R. uakarii bezeichnet, ebenfalls dieser Art zugeschrieben werden können oder ob es sich dabei um weitere neue Arten handelt, ist noch nicht abschließend zu beurteilen. In Amazonien scheint es aber etliche Populationen von verschiedenen Arten zu geben die sich in Farbe und Muster nahezu perfekt kopieren (Kopierarten nach SCHULTE, 1999). Auch das Vorkommen von sehr ähnlichen Geschwisterarten (vgl. auch LÖTTERS ET AL., 2000) erschweren eine korrekte Artbestimmung. Wie bereits CALDWELL ET AL. (1990) aufzeigen konnten verbergen sich unter der Sammelbezeichnung R. ventrimaculata noch weitere unbeschriebene Arten. Neueste Vergleiche der genetischen Daten von BROWN ET AL. (2006) liefern dafür weitere Hinweise. So konnten drei genetisch voneinander getrennte Ventrimaculatus-n abgegrenzt werden. Die drei Kladen sind nicht , können also keinen gemeinsamen Vorfahren besitzen und müssen somit auch unterschiedlichen Arten angehören. Interessant ist, das die Art R. uakarii trotz der großen äußeren Ähnlichkeiten zu R. duellmani nicht dessen nächster Verwandter zu sein scheint. Während R. duellmani mit seiner Schwesterart R. reticulata die Reticulata-Klade bildet, bildet R. uakarii eine eigene Klade die R. fantastica genetisch näher steht (BROWN ET AL., 2006)
| Bedrohungsstatus | Einklappen |
Anhang II des WA. Anhang B der EU-ArtSchVO(EG). Meldepflichtig nach BArtSchVO.
Keine Angaben nach IUCN oder CITES, da dort noch nicht als Art gelistet. Bis zur Klärung des Artstatus morphologisch ähnlicher Populationen bleibt das Verbreitungsgebiet eng umgrenzt. R. uakarii von Tahuayo-Tamshiyacu lebt aber in einem Schutzgebiet und ist damit im Moment nicht gefährdet.
Keine Angaben nach IUCN oder CITES, da dort noch nicht als Art gelistet. Bis zur Klärung des Artstatus morphologisch ähnlicher Populationen bleibt das Verbreitungsgebiet eng umgrenzt. R. uakarii von Tahuayo-Tamshiyacu lebt aber in einem Schutzgebiet und ist damit im Moment nicht gefährdet.
| Beschreibung | Einklappen |
| Größe Kleine Pfeilgiftfrosch-Art von 18-20 mm Größe. Der besitzt nach BROWN ET AL. (2006) nur eine Größe von 16,16 mm. In Gefangenschaft gehaltene adulte Tiere sind aber größer (OSTROWSKI, pers. Beobachtung). |  © 2008 Thorsten Mahn |
Das Aussehen von Ranitomeya uakarii ähnelt in vielen Farbmerkmalen dem von Ranitomeya duellmani. Auf dunklem Rücken liegen zwei in rotorange (rot bei R. duellmani). Diese verbinden sich auf dem Nasenrücken in einem umlaufenden Bogen (U-Streifen). Auf dem Rücken befindet sich in gleicher Farbe ein . Dieser reicht vom Steißbein bis zwischen die Augen. Medianstreifen und Dorsolateralstreifen sind meist durch schwarze Dorsalbänder getrennt. Im Bereich der Schnauze kann es aber auch zu einer Unterbrechung der schwarzen Zeichnung kommen. Dann findet sich auf dem Nasenrücken ein schwarzer runder Fleck. In einigen Farbmerkmalen gibt es aber auch deutliche Unterschiede zu Ranitomeya duellmani. Im Gegensatz zu R. duellmani findet sich bei R. uakarii einen in gelb, während dieser bei R. duellmani die gleiche Farbe wie die Rückenzeichnung besitzt. Weiterhin besitzt R. uakarii einen gelegenen Lateralstreifen in gelb, welcher R. duellmani fehlt. R. uakarii zeigt ein Ventralband in schwarz, das aber oft in Flecke aufgelöst ist. Die blau-schwarze Netzzeichnung (Blasenmuster) der Bauchseite zieht nicht bis auf die Flanken, wie bei R. duellmani. Ein wichtiges Unterscheidungsmerkmal ist das Vorhandensein gelber Signalflecken am Ansatz der Vorder- und Hinterbeine. Diese fehlen bei R. duellmani vollständig. Ein weiteres Unterscheidungskriterium ist die Kehlzeichnung die nach SCHULTE (1990) bei R. duellmani s.s. einen roten Signalfleck aufweist, der an seine Schwesternart R. reticulata erinnert. In den Populationen Tamshiyacu und Tahuayo findet man ein anderes Zeichnungsmuster in gelb mit schwarzer Fleckenzeichnung, welches an R. ventrimaculata erinnert. Ob es sich bei denen im Zeichnungsmuster sehr ähnlichen Populationen vom Río Sira und Río Napo (hier als R. cf. uakarii bezeichnet) ebenfalls um Varianten von R. uakarii oder um neue Arten handelt bleibt abzuwarten. Während die Variante vom Río Napo nur in der Beinfärbung (gelbes Blasennetzwerk) von R. uakarii abweicht, sind die Tiere vom Río Sira morphologisch deutlicher von R. uakarii s.s. zu unterscheiden. Sie erreichen Kopf-Rumpf-Längen von bis zu 22 mm und ihnen fehlt auch das typische in gelb und die Signalflecken von R. uakarii. Wie bei R. duellmani erreicht die blau marmorierte Bauchfärbung hier im Gegensatz zu R. uakarii die Flanken und grenzt direkt an das schwarze Lateralband. Die Population vom Río Pachitea zeigen damit eine e Zeichnung zwischen R. uakarii und R. duellmani. Es dürfte sich deshalb wohl bei dieser Population eher um eine weitere Geschwisterart innerhalb der Ventrimaculatus Gruppe handeln. In der Kehlfärbung stimmen alle hier aufgeführten Populationen überein. Dieses Merkmal haben sie jedoch mit vielen Populationen von R. ventrimaculata s.l. (im Sinne von CALDWELL ET AL., 1990) gemeinsam. Es dürfte sich bei der gelb schwarz gemusterten Signalkehle um ein eher ursprüngliches Merkmal der Ventrimaculatus-Gruppe handeln.
Text & Zeichnungen: © 2005 T. Ostrowski.
| Ranitomeya duellmani s.s. |  |  |
| Ranitomeya uakarii "Tahuayo" |  |  |
| Ranitomeya cf. uakarii "Napo" |  |  |
| Ranitomeya cf. uakarii "Pachitea" | | 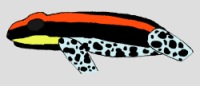 |
 Abb.: ()Dorsal- und Ventralansicht von Ranitomeya uakarii © 2006 () | Größe vergleichen: Dorsal- und Ventralbild vergleichen: |
Tab.: Messwerte ausgewählter morphologische Merkmale von Ranitomeya uakarii, Holotyp. (Datensätze mit der Kennzeichnung *) sind nicht Geschlechtsspezifisch.)
| X [mm] | σ +/- [mm] | n | Quelle | ||||
| ♂ | ♀ | ♂ | ♀ | ♂ | ♀ | ||
| 15,03 | 15,93 | 0,31 | 0,23 | 3 | 2 | BROWN ET AL., 2006 | |
| 6,28 | 6,45 | 0,03 | 0,01 | 3 | 2 | BROWN ET AL., 2006 | |
| 6,95 | 7,08 | 0,17 | 0,03 | 3 | 2 | BROWN ET AL., 2006 | |
| 12,95 | 13,16 | 0,17 | 0,46 | 3 | 2 | BROWN ET AL., 2006 | |
| 5,95 | 6,05 | 0,15 | 0,03 | 3 | 2 | BROWN ET AL., 2006 | |
| 3,69 | 3,97 | 0,09 | 0,14 | 3 | 2 | BROWN ET AL., 2006 | |
| 5,19 | 5,22 | 0,22 | 0,28 | 3 | 2 | BROWN ET AL., 2006 | |
| 5,09 | 5,06 | 0,14 | 0,11 | 3 | 2 | BROWN ET AL., 2006 | |
| 5,15 | 5,86 | 0,27 | 0,02 | 3 | 2 | BROWN ET AL., 2006 | |
| 1,33 | 1,31 | 0,04 | 0,07 | 3 | 2 | BROWN ET AL., 2006 | |
| 2,21 | 2,31 | 0,06 | 0,04 | 3 | 2 | BROWN ET AL., 2006 | |
| 0,55 | 0,60 | 0,05 | 0,02 | 3 | 2 | BROWN ET AL., 2006 | |
| 1,61 | 1,49 | 0,02 | 0,02 | 3 | 2 | BROWN ET AL., 2006 | |
| 1,11 | 1,22 | 0,09 | 0,02 | 3 | 2 | BROWN ET AL., 2006 | |
| 1,53 | 1,69 | 0,08 | 0,00 | 3 | 2 | BROWN ET AL., 2006 | |
| 0,56 | 0,60 | 0,02 | 0,02 | 3 | 2 | BROWN ET AL., 2006 | |
| *) | 1<2<4<3 | BROWN ET AL., 2006 | |||||
| 1<2<5<3<4 | BROWN ET AL., 2006 | ||||||
Variation:
BROWN ET AL. (2006) beschreiben zwei Varianten, die sich in Körpergröße und in der Breite der schwarzen Körperbänder leicht unterscheiden. Tiere vom Río Yarapa, südlich der Reserva Tamshiyacu-Tahuayo, sind etwas kleiner und besitzen breitere schwarze Streifen . Die gelben Lateral- und Ventralbänder sind außerdem intensiver gelb gefärbt als bei Tieren aus der Population am Río Tahuayo (s. Fotos rechts). Die Variation innnerhalb einer Population scheint jedoch gering zu sein. So zeigen z.B. einige wenige Nachzuchten der Tahuayo Variante, statt eines schwarzen U-Streifens um die Schnauze, einen runden Nasenfleck (OSTROWSKI, pers. Beobachtung).
Tab.: Namen, Größenangaben und Farbmusterverteilung verschiedener Populationen von Ranitomeya uakarii
| Farbmorphe | Foto | Alternativ Namen | Verbreitung | Grundfarbe | Zeichnung | Größe [mm] |
| Holotyp |  | |||||
| Napo | ||||||
| Pachitea | ||||||
| Südlichere Variante Yarapa | ||||||
| Tamshiyacu |
Wie andere Arten der Ventrimaculatus-Gruppe bis zu 10 Jahre. Meist um die 5-6 Jahre.
Geschlechtsreife:
Mit ca. 8 Monaten beginnen die ersten Männchen zu rufen. Gelege werden etwa mit 10 Monaten abgesetzt. Die ersten Gelege entwickeln sich oft nicht oder die Larven schlüpfen nicht selbstständig. Nach etwa 12 -14 Monaten werden dann „gute“ Gelege produziert und die Schlupfrate erhöht sich von 30 auf 90 % (OSTROWSKI, pers. Beobachtung)
Hautgifte:
Nicht bekannt.
| Gelege und Larven | Einklappen |
Merkmale Gelege:
Merkmale Larven:
Entwicklungszeiten:
Abb.: Ranitomeya sp. aff. uakarii beim Transport von 2 Larven
© 2008 Frank Steinmann
Ernährung:
Die Tiere legen 2-5 Eier (∅ 3,6 Eier, n = 19) mit einem Durchmesser von 1,5 mm.
Merkmale Larven:
Die Larven schlüpfen mit 12 mm Gesamtlänge (: 4mm). Im Alter von 60 Tagen messen sie etwa 27 mm (KRL: 10 mm). Wie auch bei anderen Arten der Ventrimaculatus-Gruppe, zeigen die Larven schon mit der Bildung der Hinterbeine eine beginnende Färbung am Kopf. Sie gehen mit voll entwickeltem Zeichnungsmuster in die Metamorphose. Auch die Färbung ist dann bereits erkennbar, hellt lediglich in den ersten 4-5 Tagen nach dem Landgang noch etwas auf und gewinnt an Brillanz.

Abb.: Ranitomeya sp. aff. uakarii beim Transport von 2 Larven
© 2008 Frank Steinmann
Nach etwa 12 - 15 Tagen schlüpfen die Larven aus den Eiern. Mit etwa 45 - 50 Tagen sind die Hinterbeine gut erkennbar und die Kopfzeichnung ist deutlich sichtbar, die Vorderbeine brechen nach 65 - 70 Tagen aus den Vorderbeintaschen. Die Metamorphose ist nach 70 - 75 Tagen mit der vollständigen Resorption des Schwanzes abgeschlossen. (OSTROWSKI, pers. Beobachtung; n = 14)
Ernährung:
Mit einem fein gemahlenen Gemisch aus Zierfisch-Flockenfutter , Spirulina-Algen und gefriergetrockneten Gliedertieren wie Bachflohkrebsen, Roten Mückenlarven und Artemia, konnten die Larven sehr gut aufgezogen werden. (OSTROWSKI, eigene Beobachtung)
| Verhalten (Ethologie) | Einklappen |
Allgemeines Verhalten:
Revierverhalten:
Balzverhalten:
Brutpflegeverhalten:
Abb.: Als mögliche Larvenabsatzstellen kommen Bromelien in Frage. Tamshiyacu, Peru
© 2007 Reimar David
Lautäußerung (Vokalisation):
Ruf Ranitomeya uakarii, © T. Ostrowski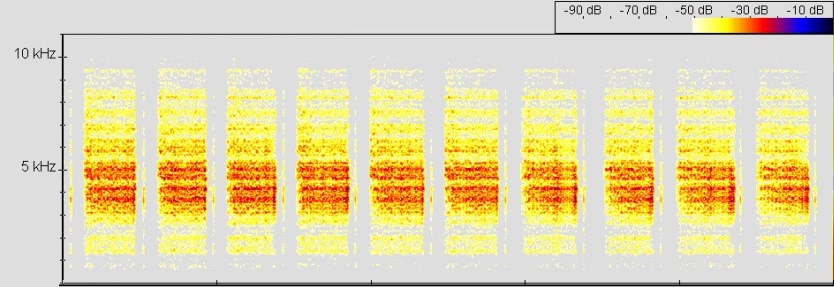
Abb.: ()Spectrogram, FFT size 512, Hanning window
Sonagramm vergleichen:
Die Tiere sind wohl ähnlich wie Ranitomeya reticulata Bewohner der Laubstreu. Laut BROWN ET AL. (2006) verbringen die Tiere die meiste Zeit am Boden, klettern aber auch gelegentlich. NIESZPOREK (pers. Komm.) berichtet, dass die Tiere im Freiland unabhängig von der Witterung den ganzen Tag aktiv sind. Er traf nur dort Tiere an, wo dicke Laubschichten vorhanden waren. R. uakarii ist ein ruhiger, scheuer Frosch. Die Tiere „tauchen“ laut NIESZPOREK (pers. Komm) im Biotop fluchtartig in der Laubstreu ab und flüchten auch unter der Laubschicht noch 1 - 2 m. Im Terrarium sind die Tiere ebenfalls recht schreckhaft und springen bei Störungen hektisch durch das Terrarium. Bei vorsichtiger Annäherung sind sie jedoch gut zu beobachten (OSTROWSKI, pers. Beobachtung).
Revierverhalten:
Über das Revierverhalten im Biotop liegen bisher keine Beobachtungen vor. Im Terrarium lassen sich gemeinsam aufgezogene Tiere gut in der Gruppe halten. Eine Zuchtgruppe von OSTROWSKI bestehend aus 2 Männchen und 3 Weibchen bewohnt ein Terrarium von 30 cm x 20 cm x 40 cm. Es wurden von beiden Männchen und allen Weibchen Paarungen mit befruchteten Gelegen beobachtet. Aggressives Verhalten konnte bisher nicht beobachtet werden. Auch der verwandte Ranitomeya ventrimaculata ist gut in der Gruppe haltbar und im Terrarium wenig aggressiv. Dennoch liegen für ihn Berichte über Aggressivität zwischen Männchen vor (SUMMERS ET AL., 1997). Im Freiland oder in größeren Terrarien könnten die Tiere also durchaus wesentlich territorialer sein, als bisher beobachtet.
Balzverhalten:
Das Balzverhalten von R. uakarii wurde von SKUDLAREK (pers. Mitteilung) und OSTROWSKI mehrmals beobachtet und dokumentiert. Vor allem in den Morgenstunden und den späten Abendstunden sind die Tiere mit der Balz beschäftigt. Das Männchen gibt bei Sichtkontakt seinen leisen Ruf ab. Dabei positioniert es sich mit aufgerichteten Vorderbeinen in Blickrichtung zum Weibchen. Reagiert das Weibchen nicht, versucht das Männchen durch staksende Bewegungen und weiteren Sichtkontakt mit dem Weibchen dieses umzustimmen. Reagiert das Weibchen auf die Rufe, so nähert sich dieses, ebenfalls aufgerichtet, schnell und zielstrebig dem Männchen. Es versucht nun das Männchen am Hinterleib zu berühren. Oft klettert es dabei auch über den Körper des Männchens. Das Männchen versucht seinerseits dem Weibchen mit dem Kopf zugewandt zu bleiben und dabei weiterhin Rufe abzugeben. Dadurch umrunden sich die Tiere mit hektisch wirkenden staksenden Bewegungen gegenseitig. Dabei kommt es immer häufiger, durch Überklettern oder Vorderbeinstreicheln seitens des Weibchens, zum Körperkontakt. Diese Phase kann bis zu 20 min dauern, bis das Männchen beginnt das Weibchen zu Laichstelle zu locken. Dazu durchbricht das Männchen nach einigen Umrundungen das Umkreisen des Weibchens und bewegt sich unter Rufen etwas zur Seite, bis das Weibchen folgt. Dann beginnt erneutes Umkreisen. Das Männchen lockt mit dem kurzzeitigen Abbrechen der Umkreisungen das Weibchen zu einem Eiablageplatz seiner Wahl (meist glatte, geschützte Flächen wie in Fotodosen oder unter Laubblättern). Auch wenn diese Ablaichstelle nur wenige cm vom Ort der Balz entfernt ist, dauert es oftmals 30 – 45 min bis die Stelle erreicht ist. Dort berühren sich die Tiere immer wieder und umkreisen sich auch. Oft verharren sie einen Moment bewegungslos Kopf an Kopf nebeneinander, wobei immer Körperkontakt gehalten wird. Bleibt das Weibchen nun ruhig neben dem Männchen sitzen, so werden dessen Rufe immer kürzer. Schließlich verstummt es und verharrt neben dem Weibchen. Verlässt das Weibchen nun ohne Eiablage den Laichort, so beginnt das Männchen wieder mit staksenden Bewegungen und verstärktem Rufen das Weibchen zu umwerben. Eine Eiablage konnte bisher nicht beobachtet werden, da trotz Erreichen des bevorzugten Laichplatzes nicht immer Gelege abgesetzt wurden, sondern die Balz oft abgebrochen wurde und erst Stunden später oder am nächsten Tag wieder fortgeführt wurde. Für Arten der Ventrimaculatus-Gruppe scheint eine solch langen Balzhandlungen die Regel zu sein, wie Beobachtungen an R. amazonica und R. variabilis zeigen (OSTROWSKI pers. Beobachtung u. ISENSEE, pers. Komm.). Auch für R. ventrimaculata sind mit 9 - 24 h Balz extrem lange Balzhandlung dokumentiert (SUMMERS ET AL., 1999 u. ZIMMERMANN & ZIMMERMANN, 1988). Offenbar ist eine gute Paar-Synchronisation für ein erfolgreiches Abschließen der Balzhandlungen bei den kleinen amazonischen Ranitomeya-Arten sehr wichtig.
Brutpflegeverhalten:

Abb.: Als mögliche Larvenabsatzstellen kommen Bromelien in Frage. Tamshiyacu, Peru
© 2007 Reimar David
Es liegen bisher nur wenige Beobachtungen aus dem Freiland vor. NIEZPOREK (pers. Komm.) konnte Larven tragende Männchen etwa 1 m über dem Boden in Bromelien beobachten. Auch BROWN ET AL. (2006) konnten Männchen beim Transport von 1-4 Larven in große
beobachten. Terrarienbeobachtungen zeigen, dass das Männchen die Gelegepflege alleine übernimmt. Es bewässert die Eier und transportiert nach dem Schlupf die Larven, oft alle gemeinsam. Diese werden dann in einer oder mehreren kleinen Wasseransammlungen abgesetzt. Die Gelege werden auf trockenen waagerechten abgestorbenen Blättern oder anderen glatten Stellen am Boden abgelegt. Dabei ist kein stehendes Wasser vorhanden, wie bei R. amazonica oder R. variabilis, die ihre Gelege am Rand von oder direkt in kleinen Phytotelmata absetzen. Die Männchen von R. uakarii scheinen die betreuten Gelege recht gut zu bewässern, um sie vor dem Austrocknen zu schützen, denn auch in Trockenperioden konnten im Biotop Männchen beim Transport der Laven beobachtet werden (PIPER, pers. Komm.).Lautäußerung (Vokalisation):
Die Tiere rufen im Terrarium vor allem in den Morgen und Abendstunden und vermehrt nach dem Nebeln oder Sprühen. Es handelt sich um einen (buzz-call). Er ähnelt dem Ruf des nah verwandten R. fantastica, besitzt aber kürzeren Pausen zwischen den (BROWN ET AL., 2006). Der Ruf ist sehr leise und kann im Freiland kaum als Anzeigeruf zur Revierabgrenzung und Partnerfindung dienen. Da die Tiere auch wenig territorial erscheinen und nur bei Sichtkontakt rufen, erscheint es wahrscheinlicher, dass es sich um einen reinen Nahbereich-Werberuf (close-range-mating-call) handelt. Eine besteht aus gepulsten Noten von 295 – 453 ms Dauer (Ø = 401 ms; n = 12). Die Noten sind durch Intervalle von 630 – 750 ms (Ø = 679 ms; n = 12) Dauer getrennt. Die beträgt im Mittel 2,03 Noten/s. Eine Sequenz beinhaltet durchschnittlich etwa 25 Noten und ist etwa 12 Sekunden lang (siehe auch Abb: Gesamtes Sonagramm). Die Noten ließen sich mit dem genutzen Programm (BatSoundPro) nicht in Einzelpulse auflösen. Es liegen somit keine vor und die überlagern sich teilweise. Typisch für eine Note von D. uakarii scheint jedoch ein gut getrennter Einzelpuls am Ende der Note. Dieser Einzelpuls hat eine Dauer von 10,4 – 18,3 ms (Ø = 14 ms; n = 12). Von dem Hauptfeld der Pulse wird er durch ein langes Intervall von 28,7 – 51,5 ms (Ø = 39,8 ms; n = 12) Dauer getrennt (s. Abb.:).
Ruf Ranitomeya uakarii, © T. Ostrowski
Abb.: ()Spectrogram, FFT size 512, Hanning window
Sonagramm vergleichen:
| Lebensraum | Einklappen |
Typenfundort der Erstbeschreibung
Daten von OpenStreetMap - Veröffentlicht unter CC-BY-SA 2.0
Biotop:
Klima:
"...upstream Quebrada Blanco in Tamshiyacu-Tahuayo Reserve,
Departamento Loreto, Peru
(4º11´21.88´´ S, 73º6´15.66´´ W), Elevation: 140 m." - BROWN ET AL., 2006.
(4º11´21.88´´ S, 73º6´15.66´´ W), Elevation: 140 m." - BROWN ET AL., 2006.
Verbreitung: | 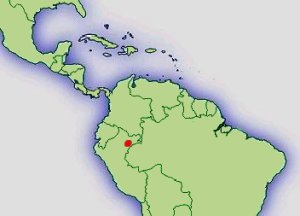 Abb.: ()Verbreitungsgebiet Ranitomeya uakarii © 2008 () |
Biotop:
Primärregenwald zwischen den Flussläufen. Nicht im Überschwemmungswald! NIESZPOREK (pers. Komm.) berichtet von der Population „Tahuayo“ in der Nähe von Iquitos. Die Tiere leben in der Laubschicht in Bodennähe. Sie klettern kaum und wurden bis max. 1 m Höhe über dem Boden angetroffen Die Tiere fanden sich an Stellen, wo genügend dicke Laubdecken vorhanden waren, zum Beispiel im Stammbasenbereich oder an gefallenen Stämmen von großen Urwaldriesen. Beim Fehlen einer dicken Laubstreu waren auch keine R. uakarii nachzuweisen.
 Abb.: Natürlicher Lebensraum von Ranitomeya uakarii. Tamshiyacu, Peru © 2007 Reimar David |
 Abb.: Bevorzugt halten sich Ranitomeya uakarii in der Laubschicht auf. Tamshiyacu, Peru © 2007 Reimar David |
 Abb.: Orchideen und Bromelien im Biotop von Ranitomeya uakarii. Tamshiyacu, Peru © 2007 Reimar David |
 Abb.: Weitere Pflanzen im Biotop von Ranitomeya uakarii. Tamshiyacu, Peru © 2007 Reimar David |
 Abb.: Ranitomeya uakarii bewohnt vor allem die Laub- und Krautschicht der tropischen Sekundärwälder entlang des oberen Río Amazonas. Departamento Loreto, Peru. © 2008 Thomas Ostrowski |
 Abb.: Waldboden am Río Amazonas. Biotop von Ranitomeya uakarii. Departamento Loreto, Peru. © 2008 Thomas Ostrowski |
 Abb.: Als Brutpflanzen werden von Ranitomeya uakarii verschiedene Bromelien (hier Guzmania sp.) in den höheren lichtdurchfluteten Etagen des Waldes genutzt. Departamento Loreto, Peru. © 2008 Thomas Ostrowski |
 Abb.: Verschiedene Guzmania-Arten kommen im Biotop in der Nähe des Río Tamshiyacu vor und werden von Ranitomeya uakarii als potentielle Brutpflanzen genutzt. Departamento Loreto, Peru. © 2008 Thomas Ostrowski |
 Abb.: Eine blühende Guzmania sp., Brutpflanze von Ranitomeya uakarii. Departamento Loreto, Peru. © 2008 Thomas Ostrowski |
Klima:
Die Biotope liegen alle in feuchtheißen Tieflandregenwäldern mit Tageszeitenklima. Ganzjährig herrschen recht konstante Temperaturen. Die Schwankungen der Temperatur im Jahresmittel fallen mit 1 - 2 ° C geringer aus als die Schwankungen zwischen Tages- und Nachttemperaturen. Aber auch diese liegen im Tieflandregenwald des Amazonasgebietes nur bei 3 - 4 ° C. Am Waldboden lässt sich kaum noch eine Temperaturdifferenz nachweisen und schon dicht unter der Laubschicht lassen sich Temperaturen messen, die in etwa denen des Jahresmittels entsprechen (WALTER & BRECKLE, 1999). Die Temperaturen innerhalb der Laubschicht dürften deshalb in etwa dem Jahresmittel von Iquitos mit 26 °C entsprechen. Das Klima des Amazonasbeckens ist auch in Bezug auf die Niederschläge recht konstant und ganzjährig humid. In den Monaten Juni bis Oktober gehen die Niederschläge zwar etwas zrück, aber es gibt keine ausgeprägte Trockenperiode.
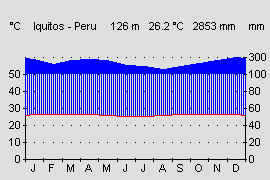 Klimadiagramm des amazonischen Tieflandes am Beispiel Iquitos. Quelle: www.klimadiagramme.de |
| Haltung im Terrarium | Einklappen |
Terrarium/Einrichtung:
Temperaturen:
Luftfeuchte:
Ernährung:
Besatz:
Tipps zur Zucht:
Varianten in Terrarienhaltung:
ab 40 cm x 40 cm x 40cm. Einrichtung mit dichter Bepflanzung und Laubschicht am Boden. Waagerechte Fotodosen als und Versteckmöglichkeit anbieten. Eine automatische Beregnung und Nebelanlage ist empfehlenswert
Temperaturen:
Als Bewohner des amazonischen Tieflandregenwaldes benötigen die Tiere Temperaturen von 25-27° C.
Nachtabsenkung von 2-4° C empfohlen.
Nachtabsenkung von 2-4° C empfohlen.
Luftfeuchte:
75-85%, zur Mittagszeit bis 70%, morgens und abends 100% (Nebel)
Ernährung:
Übliche kleine Futtertiere wie , , bzw. Heimchen, frisch geschlüpfte n, (Mikrokäfer), und fein gesiebtes . Für Jungtiere sind in den ersten zwei Wochen Springschwänze unabdingbar. Nur Jungfrösche die aus sehr großen Larven metamorphisiert sind, können manchmal auch schon sofort die Kleine Obstfliege bewältigen. Für sollten Futtertiere wie Grillen und Obstfliegen regelmäßig 1-2 mal die Woche mit einem guten Vitaminpräparat eingestäubt werden (z.B. Amivit A nach der orginal BIRKHAHN-Rezeptur). Futtertiere für Jungtiere sollten die ersten 4 Wochen täglich bestäubt werden. Angebrochene Vitaminpräparate im Kühlschrank aufbewahren. Obstfliegen lassen sich vor dem Verfüttern gut mit flüssigen Vitaminpräparaten (z.B. Sanostol, Multibionta) anfüttern und so ernährungsphysiologisch aufwerten. Bestäubte Futtertiere sollten im Terrarium auf auswechselbaren Schalen angeboten werden. Zurückbleibende Vitaminpulverreste können so keinen Bakterienherd auf dem Terrariumboden bilden. In kleinen Schalen im Terrarium ausgelegte Obststücke (z.B. Bananenscheiben) sind gute Lockstellen für Obstfliegen und werden von den Fröschen bald als Futterplätze akzeptiert. Für eine ausreichende Vitaminversorgung der Futtertiere durch diese Lockstellen dürfte die Verweildauer der Futtertiere jedoch zu gering sein, so dass dennoch zusätlich vitaminisiert werden sollte. Angebotene Futterschalen sollten aus hygienischen Gründen alle 2-3 Tage gereinigt werden. Springschwänze lassen sich gut auf ausgelegten Xaxim-Stückchen konzentrieren, indem man diese mit kleinen! Mengen Trockenhefe bestreut. Auch hier lernen die Frösche schnell die Bedeutung des Futterplatzes.
Besatz:
Beste Ergebnisse mit 1,1. Gruppenhaltung möglich, z.B. 2,3 in 50 cm x 50 cm x 50 cm.
Tipps zur Zucht:
Die Tiere laichen in waagerecht angebrachten schwarzen Fotodosen.
Früh entfernte, separat gezeitigte Gelege entwickeln sich besser, als im Terrarium belassene Gelege. Einzelhaltung der kannibalisch veranlagten Quappen ist nötig. Javamoos und eine kleine Wasserschnecke bei der Quappe beugen verdorbenem Futter und Keimbelastung des Wassers vor. Wassertemperatur bei 24°C, Nachtabsenkung um 2-5 ° C empfohlen. Die Aufzucht der Jungtiere kann in Gruppen erfolgen. Die Jungtiere wachsen sehr schnell. (OSTROWSKI, pers. Beobachtung).
Varianten in Terrarienhaltung:
| Holotyp |  |
| Napo |  |
| Pachitea |  |
| Südlichere Variante Yarapa |  |
| Tamshiyacu |  |
Daten errechnet aus von Nutzern gemeldeten Fröschen (Home->Meine Frösche) und den aktuellen Inseraten.
stand 10.03.2026 01:10 Uhr
stand 10.03.2026 01:10 Uhr
| Autoren dieser Seite: | Einklappen |
| Referenzen | Einklappen |
BROWN, J.L. , R. SCHULTE, & K. SUMMERS (2006): A new species of Dendrobates (Anura: Dendrobatidae)
XXfrom the Amazonian lowlands in Perú - Zootaxa 1152: 45–58 (PDF)
CALDWELL, J.P. & C.W. MYERS (1990): A new poison frog from Amazonian Brazil, with further
XX revision of the quinquevittatus Group of Dendrobates. - Am.Mus.Nov. 2988: 1-21. (PDF)
CITES (2005): Convention on International Trade of Endangered Species of Wild Fauna and Flora.
XXOnlineversion: 11.08.05. Electronic Database accessible at http://www.cites.org/index.html
FROST, D.R. (2007): Amphibian Species of the World: an Online Reference. Version 5.0 (1.02.2007).
XXElectronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.php
XX Am.Mus.Nat.Hist. New York, USA.
GRANT, T . , FROST, D. R. , CALDWELL, J. P. , GAGLIARDO, R. , HADDAD, C. F. B. , KOK, P. J. R. ,
XXMEANS, D.B. , NOONAN, B. P. , SCHARGEL, W. E. & W.C. WHEELER (2006): Phylogenetic
XXsystematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae).
XX - Bulletin of the Am.Mus.Nat.Hist. 299. (PDF)
IUCN, Conservation International, and NatureServe. (2009): Global Amphibian Assessment.
XXwww.globalamphibians.org. Downloaded on 15 October 2004.
LÖTTERS, S. & M. VENCES (2000): Bemerkungen zur Nomenklatur und Taxonomie peruanischer
XXPfeilgiftfrösche. - Salamandra 36 (4): 247-260. (PDF)
SCHULTE, R. (1999): Pfeilgiftfrösche - Artenteil Peru. - Nikola Verlag, Stuttgart.
SUMMERS, K. & W. AMOS (1997): Behavioral, ecological, and molecular genetic analyses of reproduction
XXstrategies in the Amazonian dart-poison frog, Dendrobates ventrimaculatus.
XX- Behav Ecol 8(3): 260-267.
WALTER, H. & S. BRECKLE (1999): Vegetationen und Klimazonen. 7. Auflage. - Ulmer, Stuttgart.
ZIMMERMANN, E. & H. ZIMMERMANN (1984): Durch Nachzucht erhalten: Baumsteigerfrösche
XXDendrobates quinquevittatus und D. reticulatus. – Aquarien Mag. 18(1): 35-41.
XXfrom the Amazonian lowlands in Perú - Zootaxa 1152: 45–58 (PDF)
CALDWELL, J.P. & C.W. MYERS (1990): A new poison frog from Amazonian Brazil, with further
XX revision of the quinquevittatus Group of Dendrobates. - Am.Mus.Nov. 2988: 1-21. (PDF)
CITES (2005): Convention on International Trade of Endangered Species of Wild Fauna and Flora.
XXOnlineversion: 11.08.05. Electronic Database accessible at http://www.cites.org/index.html
FROST, D.R. (2007): Amphibian Species of the World: an Online Reference. Version 5.0 (1.02.2007).
XXElectronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.php
XX Am.Mus.Nat.Hist. New York, USA.
GRANT, T . , FROST, D. R. , CALDWELL, J. P. , GAGLIARDO, R. , HADDAD, C. F. B. , KOK, P. J. R. ,
XXMEANS, D.B. , NOONAN, B. P. , SCHARGEL, W. E. & W.C. WHEELER (2006): Phylogenetic
XXsystematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae).
XX - Bulletin of the Am.Mus.Nat.Hist. 299. (PDF)
IUCN, Conservation International, and NatureServe. (2009): Global Amphibian Assessment.
XXwww.globalamphibians.org. Downloaded on 15 October 2004.
LÖTTERS, S. & M. VENCES (2000): Bemerkungen zur Nomenklatur und Taxonomie peruanischer
XXPfeilgiftfrösche. - Salamandra 36 (4): 247-260. (PDF)
SCHULTE, R. (1999): Pfeilgiftfrösche - Artenteil Peru. - Nikola Verlag, Stuttgart.
SUMMERS, K. & W. AMOS (1997): Behavioral, ecological, and molecular genetic analyses of reproduction
XXstrategies in the Amazonian dart-poison frog, Dendrobates ventrimaculatus.
XX- Behav Ecol 8(3): 260-267.
WALTER, H. & S. BRECKLE (1999): Vegetationen und Klimazonen. 7. Auflage. - Ulmer, Stuttgart.
ZIMMERMANN, E. & H. ZIMMERMANN (1984): Durch Nachzucht erhalten: Baumsteigerfrösche
XXDendrobates quinquevittatus und D. reticulatus. – Aquarien Mag. 18(1): 35-41.
| Zitierung: | Einklappen |
OSTROWSKI, T. & T. MAHN (2026): Artbeschreibung Ranitomeya uakarii.
- Dendrobase.de - Eine Online-Datenbank der Familie Dendrobatidae (Anura).
https://www.dendrobase.de/html/D_ranitomeya_uakarii.html; Stand: März 2026
- Dendrobase.de - Eine Online-Datenbank der Familie Dendrobatidae (Anura).
https://www.dendrobase.de/html/D_ranitomeya_uakarii.html; Stand: März 2026
| Link für Website Betreiber: | Einklappen |
Webmaster dürfen Links von Ihren Seiten zur DendroBase erstellen, wenn sie folgende Kriterien erfüllen:
- Die Website enthält weder illegale, rassistische oder sexistische Inhalte
- Das Thema der Website bezieht sich auf Frösche
- Der Link genau wie folgend angegeben verwendet wird:
<a href="https://www.dendrobase.de/html/D_ranitomeya_uakarii.html" target="_new">Ranitomeya uakarii (DendroBase.de)</a>
- Die Website enthält weder illegale, rassistische oder sexistische Inhalte
- Das Thema der Website bezieht sich auf Frösche
- Der Link genau wie folgend angegeben verwendet wird:
<a href="https://www.dendrobase.de/html/D_ranitomeya_uakarii.html" target="_new">Ranitomeya uakarii (DendroBase.de)</a>
| Fotos | Einklappen |

Ranitomeya ()uakarii
"Tahuayo"
© 2006 ()

Ranitomeya ()uakarii
"Tahuayo"
© 2006 ()

Ranitomeya ()uakarii
"Tahuayo"
© 2006 Joachim Bitz

Ranitomeya ()uakarii
"Tahuayo"
© 2004 Jan Stìnièka

Ranitomeya ()uakarii
"Tamshiyacu"
© 2008 ()

Ranitomeya ()uakarii
"Yarapa"
© 2005 Wojtek Nieszporek 2005 Uwe Nicolaus

Ranitomeya ()uakarii
"Yarapa"
© 2005 Wojtek Nieszporek 2005 Uwe Nicolaus

Ranitomeya ()uakarii
"Yarapa"
© 2005 Wojtek Nieszporek 2005 Uwe Nicolaus

Ranitomeya ()cf. uakarii
"Napo"
© 2005 ()

Ranitomeya ()cf. uakarii
"Napo"
© 2005 ()

Ranitomeya ()uakarii "Napo"
Weibchen mit abweichendem Zeichnungsmuster
© 2006 K. U. Stolarski

Ranitomeya ()cf. uakarii
"Pachitea"
© 2004 Andreas Toebe
DendroBase.de Version 5 | Impressum | Datenschutzerklärung