| Login | Einklappen |
Startseite
Artbeschreibungen
Allobates
Anomaloglossus
Rheobates
Aromobates
Mannophryne
Ameerega
Colostethus
Epipedobates
Silverstoneia
Andinobates
Adelphobates
Dendrobates
Excidobates
Minyobates
Oophaga
Phyllobates
Ranitomeya
Hyloxalus
Foren
Inserate
Reportagen
Service & Links
Einstellungen
| Fotos von Nutzern | Einklappen |
| Gattung Oophaga: | Einklappen |
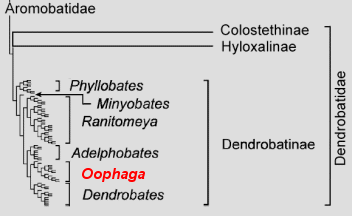
Die Gattung Oophaga wurde bereits 1994 durch BAUER von der Gattung Dendrobates WAGLER, 1830 abgetrennt. Dieser taxonomische Akt wurde jedoch mangels nachvollziehbar wissenschaftlicher Begründung nicht anerkannt und das Oophaga BAUER 1994 wurde als Synonym zu Dendrobates WAGLER 1830 angesehen. Im Zuge der umfangreichen Revision der Familie Dendrobatidae trennten GRANT ET AL. (2006) die Gattung Dendrobates aufgrund genetischer, ethologischer und morphologischer Gemeinsamkeiten erneut in mehrere Gattungen auf. Jede der vorgeschlagenen Taxa (Gattungen) bildet nach den vorliegenden Daten eine klar abgrenzbare e Gruppe im Verwandtschaftsbaum der Familie Dendrobatidae. Auch für die Arten aus der sogenannten Histrionicus-Gruppe ( MYERS & DALY 1984) von Dendrobates ergibt sich eine klar abgrenzbare . Die Beschreibung von BAUER enthielt zwar nicht genügend stichhaltige Belege für eine gesicherte Abtrennung, genügte aber in ihrer Art und Weise den internationalen Nomenklatur-Regeln des International Code of Zoological Nomenclature (abgekürzt ICZN). Somit war für die monophyletische Gruppe nach GRANT ET AL. (2006) bereits ein älterer Gattungsname verfügbar: Oophaga BAUER 1994. Dieser hat nach den Richtlinien der ICZN (2001) Vorrang vor neueren Gattungsnamen und wurde deshalb von GRANT ET AL. revalidiert.
Typusart der Gattung ist Dendrobates pumilio SCHMIDT 1857. Der Gattungsname Oophaga entstammt dem griechischen oophag (= Ei fressend) und bezieht sich auf die hochspezialisierte Brutpflege (Larvenfütterung mit n) innerhalb der Gattung. Innerhalb der Familie Dendrobatidae steht die Gattung als eigenständige Klade in der Unterfamilie Dendrobatinae COPE 1865 gemeinsam mit der Schwestergattung Dendrobates WAGLER 1830 neben den übrigen Gattungen Adelphobates GRANT ET AL. 2006, Minyobates MYERS, 1982, Phyllobates DUMERIL & BIBRON 1841 und Ranitomeya BAUER, 1988. Die Gattung enthält zurzeit 9 gültige Arten: Oophaga arborea, O. granulifera, O. histrionica, O. lehmanni, O. occultator, O. pumilio, O. speciosa, O. sylvatica und O. vicentei * (sensu FROST, 2007). Die Abgrenzung zu den anderen Gattungen erfolgte aufgrund er genetischer, er und er Merkmale. Die genetisch monophyletische Klade lässt sich von allen anderen Gattungen durch weitere Merkmale abgrenzen. Morphologisch vor allem durch einige Larvencharakteristika wie das vergrößerte Mundfeld, den stark ausgeprägten und vergrößerte n (an angepasstes ). Ethologisch kennzeichnet sich die Gattung durch den gemeinsamen (chirp-call) und die weibliche Brutpflege mit spezialisierter Eifütterung (e Oophagie). GRANT ET AL. (2006) führen einige weitere Merkmale an, die allerdings nicht immer eindeutig sind und teilweise auch nur e Merkmale der Unterfamilie Dendrobatinae darstellen. Viele der von GRANT ET AL. für die Abgrenzung verwendetet Charakteristika dienten bereits SILVERSTONE (1975) und MYERS & DALY (1984) dazu die Vertreter der Gattung Oophaga (sensu GRANT ET AL.) von anderen Vertretern der Gattung Dendrobates abzugrenzen. Diese Abgrenzung fand bisher Ausdruck in der Einteilung der Gattung Dendrobates in Untergruppen (Histrionicus- und Pumilio-Gruppe sensu SILVERSTONE, 1975 und Histrionicus-Gruppe sensu MYERS & DALY, 1984). Die Abgrenzungskriterien sind also nichts grundlegend Neues. Die Aufspaltung der Gattung ist damit nur eine konsequentere Auslegung altbekannter Kriterien, die zusätzlich auch den genetischen Daten eine stärkere Gewichtung beimisst.
Typusart der Gattung ist Dendrobates pumilio SCHMIDT 1857. Der Gattungsname Oophaga entstammt dem griechischen oophag (= Ei fressend) und bezieht sich auf die hochspezialisierte Brutpflege (Larvenfütterung mit n) innerhalb der Gattung. Innerhalb der Familie Dendrobatidae steht die Gattung als eigenständige Klade in der Unterfamilie Dendrobatinae COPE 1865 gemeinsam mit der Schwestergattung Dendrobates WAGLER 1830 neben den übrigen Gattungen Adelphobates GRANT ET AL. 2006, Minyobates MYERS, 1982, Phyllobates DUMERIL & BIBRON 1841 und Ranitomeya BAUER, 1988. Die Gattung enthält zurzeit 9 gültige Arten: Oophaga arborea, O. granulifera, O. histrionica, O. lehmanni, O. occultator, O. pumilio, O. speciosa, O. sylvatica und O. vicentei * (sensu FROST, 2007). Die Abgrenzung zu den anderen Gattungen erfolgte aufgrund er genetischer, er und er Merkmale. Die genetisch monophyletische Klade lässt sich von allen anderen Gattungen durch weitere Merkmale abgrenzen. Morphologisch vor allem durch einige Larvencharakteristika wie das vergrößerte Mundfeld, den stark ausgeprägten und vergrößerte n (an angepasstes ). Ethologisch kennzeichnet sich die Gattung durch den gemeinsamen (chirp-call) und die weibliche Brutpflege mit spezialisierter Eifütterung (e Oophagie). GRANT ET AL. (2006) führen einige weitere Merkmale an, die allerdings nicht immer eindeutig sind und teilweise auch nur e Merkmale der Unterfamilie Dendrobatinae darstellen. Viele der von GRANT ET AL. für die Abgrenzung verwendetet Charakteristika dienten bereits SILVERSTONE (1975) und MYERS & DALY (1984) dazu die Vertreter der Gattung Oophaga (sensu GRANT ET AL.) von anderen Vertretern der Gattung Dendrobates abzugrenzen. Diese Abgrenzung fand bisher Ausdruck in der Einteilung der Gattung Dendrobates in Untergruppen (Histrionicus- und Pumilio-Gruppe sensu SILVERSTONE, 1975 und Histrionicus-Gruppe sensu MYERS & DALY, 1984). Die Abgrenzungskriterien sind also nichts grundlegend Neues. Die Aufspaltung der Gattung ist damit nur eine konsequentere Auslegung altbekannter Kriterien, die zusätzlich auch den genetischen Daten eine stärkere Gewichtung beimisst.
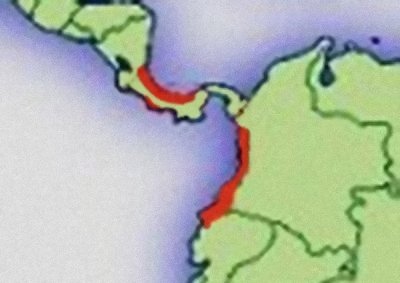
Abb.: Verbreitungsgebiet der Gattung Oophaga mit Verbreitungslücke im östlichen Panama. © 2006 Thorsten Mahn
Die Gattung Oophaga ist ein typisch westandines bzw. zentralamerikanisches Faunenelement. Die Vertreter der Gattung finden sich nur in Mittelamerika und im nordwestlichen Südamerika westlich der Andenkette (westliches Kolumbien und nordwestliches Ecuador). Das Verbreitungsgebiet deckt sich auffallend genau mit dem Verbreitungsgebiet der Gattung Phyllobates (MYERS & DALY, 1983). Beide Gattungen zeigen sogar die gleiche Verbreitungslücke im östlichen Panama. Offenbar haben beide Gattungen die gleiche biogeographische Verbreitungsgeschichte. Die Gattungen entstanden vermutlich nach der Auffaltung der Anden vor etwa 10 Mio. Jahren im Nordwesten Südamerikas (SILVERSTONE, 1975). Von hier erfolgte die Einwanderung auf die neu entstandene mittelamerikanische Landbrücke vor ca. 3,5 Mio. Jahren (SAVAGE, 2002). Zwei relativ früh eingewanderte Linien von Oophaga und Phyllobates wurden vermutlich auf der pazifischen Seite im Laufe der Erdgeschichte durch e und e Klimaveränderungen inselartig eingeschlossen. Die Art Oophaga granulifera ist vermutlich aus dieser frühen Linie hervorgegangen und ist heute die ursprünglichste Vertreterin der Gattung Oophaga. Sie zeigt sowohl mit kolumbianischen als auch mittelamerikanischen Arten nur eine entfernte Verwandtschaft. Heute sind die Arten O. granulifera und Phyllobates vittatus in Mittelamerika im Gebiet der Halbinsel Osa von allen anderen Vertretern ihrer Gattungen klimatisch durch Trockenwaldgebiete im Nordwesten und Südosten und die Hochgebirgskette im Norden abgetrennt und stellen auf der Pazifikseite in Mittelamerika ein Reliktvorkommen dar. Eine weitere Einwanderungswelle aus Südamerika folgte vermutlich der karibischen Küstenlinie. Daraus gingen z.B. die Arten O. pumilio und O. arborea hervor, die laut HAGEMANN (2005) näher mit der kolumbianischen O. histrionica verwandt sind.
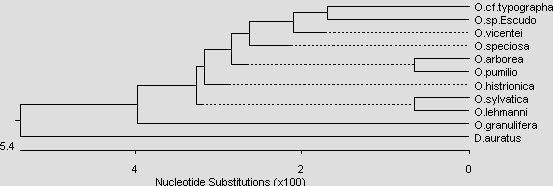
Abb.: Stammbaumartige Darstellung der Verwandtschaftsverhältnisse der Gattung Oophaga. Neighbour Joining Tree basierend auf 16S rRNA-Sequenzen. © 2007 Sabine Hagemann
Die Verbreitungslücke der Gattungen Oophaga und Phyllobates scheint ebenfalls auf glaziale und interglaziale Ereignisse zurückzuführen zu sein. Im Osten Panamas ist der sehr flach und lag vermutlich in den Zwischeneiszeiten unterhalb des Meeresspiegels. Auf dem Höhepunkt der Eiszeiten scheinen die flachen Ebenen aufgrund des trockeneren Klimas eher savannenartigen Charakter gehabt zu haben (SILVERSTONE, 1975). Diese klimatischen Schwankungen haben eine Verbreitung nur in erdgeschichtlich kurzen Zeiträumen zugelassen, in denen die Landbrücke im Bereich des Darien eine geschlossene Bewaldung aufwies. Nach der letzten Eiszeit hat es vermutlich nur der anpassungsfähige Dendrobates auratus geschafft diese Gebiete wieder erfolgreich zu besiedeln. Vertreter der Gattungen Oophaga und Phyllobates und der Minutus-Gruppe von Ranitomeya finden sich nicht im östlichen Darien sondern erst wieder östlich im Choco-Gebiet Kolumbiens bzw. westlich in Zentralpanama.
*Aufgrund des weiblichen Wortstammes der Gattung ergaben sich in neuer Kombination mit Oophaga (f) auch bei den ursprünglichen Artnamen einige Änderungen hinsichtlich der geschlechtsbestimmenden Suffixe (Wortendungen) von –us (m) nach –a (f). So wurde z.B. aus Dendroates histrionicus (m) das Taxon Oophaga histrionica (f)
| Referenzen | Einklappen |
BAUER, L. (1994): New names in the family Dendrobatidae (Anura, Amphibia). - RIPA Fall: 1–6.
BROWN & TWOMEY (2006): Dendrobatid systematics - Dendrobates.org - Ecology, evolution, and
XXconservation of poison frogs. http://www.dendrobates.org/systematics.html (Stand. 22.10.2006)
FROST, D.R. (2007): Amphibian Species of the World: an Online Reference. Version 5.0 (1.02.2007).
XXElectronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.php
XX Am.Mus.Nat.Hist. New York, USA.
GRANT, T . , FROST, D. R. , CALDWELL, J. P. , GAGLIARDO, R. , HADDAD, C. F. B. , KOK, P. J. R. ,
XXMEANS, D.B. , NOONAN, B. P. , SCHARGEL, W. E. & W.C. WHEELER (2006): Phylogenetic
XXsystematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae).
XX - Bulletin of the Am.Mus.Nat.Hist. 299. (PDF)
HAGEMANN, S. (2005): Genetische Variation beim Erdbeerfröschen (Dendrobates pumilio) in Costa Rica
XXund Panama. – Diplomarbeit, 2005. Institut für Zoologie, Tierärztlichen Hochschule Hannover.
ICZN (2001): International Code of Zoological Nomenclature. - London: International Trust for Zoological
XXNomenclature. (Electronic data accessible at: http://www.iczn.org/)
MYERS, C.W. DALY, J.W. & MARTINEZ, V. (1984): An arboreal poison frog (Dendrobates) from
XXwestern Panama.- Am.Mus.Nov. 2783: 1-20.
SAVAGE, J.M. (2002): The Amphibians and Reptiles of Costa Rica. A Herpetofauna between two
XXContinents, between two Seas. - The University of Chigago Press, Chigago.
SILVERSTONE, P.A. (1975): A Revision of the Poison-Arrow-Frogs of the Genus Dendrobates Wagler.
XX- NatHist.Mus. Los Angeles County, Sci.Bull. 21: 1-55. (PDF)
BROWN & TWOMEY (2006): Dendrobatid systematics - Dendrobates.org - Ecology, evolution, and
XXconservation of poison frogs. http://www.dendrobates.org/systematics.html (Stand. 22.10.2006)
FROST, D.R. (2007): Amphibian Species of the World: an Online Reference. Version 5.0 (1.02.2007).
XXElectronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.php
XX Am.Mus.Nat.Hist. New York, USA.
GRANT, T . , FROST, D. R. , CALDWELL, J. P. , GAGLIARDO, R. , HADDAD, C. F. B. , KOK, P. J. R. ,
XXMEANS, D.B. , NOONAN, B. P. , SCHARGEL, W. E. & W.C. WHEELER (2006): Phylogenetic
XXsystematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae).
XX - Bulletin of the Am.Mus.Nat.Hist. 299. (PDF)
HAGEMANN, S. (2005): Genetische Variation beim Erdbeerfröschen (Dendrobates pumilio) in Costa Rica
XXund Panama. – Diplomarbeit, 2005. Institut für Zoologie, Tierärztlichen Hochschule Hannover.
ICZN (2001): International Code of Zoological Nomenclature. - London: International Trust for Zoological
XXNomenclature. (Electronic data accessible at: http://www.iczn.org/)
MYERS, C.W. DALY, J.W. & MARTINEZ, V. (1984): An arboreal poison frog (Dendrobates) from
XXwestern Panama.- Am.Mus.Nov. 2783: 1-20.
SAVAGE, J.M. (2002): The Amphibians and Reptiles of Costa Rica. A Herpetofauna between two
XXContinents, between two Seas. - The University of Chigago Press, Chigago.
SILVERSTONE, P.A. (1975): A Revision of the Poison-Arrow-Frogs of the Genus Dendrobates Wagler.
XX- NatHist.Mus. Los Angeles County, Sci.Bull. 21: 1-55. (PDF)
| Fotos | Einklappen |

Oophaga ()arborea
Fotomontage
nach MYERS ET AL. (1984)
© 2007 ()

Oophaga ()granulifera
"Quepos"
© 2005 ()

Oophaga ()histrionica
"Buenaventura"
Departamento Valle de Cauca
© 2009 ()

Oophaga ()lehmanni
Departamento Valle de Cauca
© 2011 ()

Oophaga ()occultator
© 1999 ()chiller (?)
zur Verfügung gestellt von Stefan Lötters

Oophaga ()pumilio "Siquirres"
© 2006 Holger Birkhahn

Oophaga ()spec. "Escudo"
"Isla Escudo"
(Nordseite)
© 2006 ()

Oophaga ()cf. typographa
"Fortuna"
(Pueblo Nuevo)
© 2004 Jan Karsch

Oophaga ()speciosa (?)
© 2005 Ian Rogers

Oophaga ()sylvatica s.s.
"Santo Domingo"
© 2007 Marcus Bartelds

Oophaga ()vicentei
"El Copé"
© 2003 Holger Birkhahn
DendroBase.de Version 5 | Impressum | Datenschutzerklärung