| Login | Einklappen |
Startseite
Artbeschreibungen
Allobates
Anomaloglossus
Rheobates
Aromobates
Mannophryne
Ameerega
Colostethus
Epipedobates
Silverstoneia
Andinobates
Adelphobates
Dendrobates
Excidobates
Minyobates
Oophaga
Phyllobates
Ranitomeya
Hyloxalus
Foren
Inserate
Reportagen
Service & Links
Einstellungen
| Fotos von Nutzern | Einklappen |
| Nomenklatur | Einklappen |
wissenschaftl. Name: Andinobates claudiae (JUNGFER, LÖTTERS & JÖRGENS, 2000)
Etymologie:
Synonyme:
Ranitomeya claudiae (GRANT, FROST, CALDWELL, GAGLIARDO, HADDAD, KOK, MEANS, NOONAN, SCHARGEL & WHEELER, 2006)
Dendrobates claudiae JUNGFER, LÖTTERS & JÖRGENS, 2000
Dendrobates mimulus BURTON, 1998. Nomen nudum.
sensu FROST 2007
englischer Name: Bocas Poisenfrog
deutscher Name: Bocas Baumsteiger
Etymologie:
Das claudiae ist ein sname zu Ehren von CLAUDIA NOEL VLASIMSKY (JUNGFER ET AL., 2000). Sie und ihr Vater ermöglichten die Beschreibung im Rahmen einer Namenspatenschaft (BIOPAT, Katalog Nr.: 97).
Synonyme:
Ranitomeya claudiae (GRANT, FROST, CALDWELL, GAGLIARDO, HADDAD, KOK, MEANS, NOONAN, SCHARGEL & WHEELER, 2006)
Dendrobates claudiae JUNGFER, LÖTTERS & JÖRGENS, 2000
Dendrobates mimulus BURTON, 1998. Nomen nudum.
sensu FROST 2007
englischer Name: Bocas Poisenfrog
deutscher Name: Bocas Baumsteiger
| Systematik | Einklappen |
Klassifizierung:
Amphibia->Anura->Dendrobatoidea->Dendrobatidae->Dendrobatinae->Andinobates->Andinobates claudiae (JUNGFER, LÖTTERS & JÖRGENS, 2000)
Gruppe:
Anmerkung zur Systematik:
Amphibia->Anura->Dendrobatoidea->Dendrobatidae->Dendrobatinae->Andinobates->Andinobates claudiae (JUNGFER, LÖTTERS & JÖRGENS, 2000)
Gruppe:
sensu GRANT ET AL. (2006)
Anmerkung zur Systematik:
Lange Zeit wurden die Populationen der panamaischen Provinz Bocas del Toro dem Ranitomeya minuta zugeordet. Die Art R. minuta hat ihr westlichstes Verbreitungsgebiet jedoch in Zentralpanama in der Provinz Veraguas.
| Bedrohungsstatus | Einklappen |
Anhang II des WA. Anhang B der EU-ArtSchVO(EG). Meldepflichtig nach BArtSchVO.
In der Roten Liste (IUCN, 2004) als uneinstufbar (DD, Data Deficient) angegeben. Nur wenige Informationen über Verbreitung und Populationsgröße liegen vor. Ein Populationstrend ist derzeit nicht einschätzbar. Da die Inseln des Bocas Archipels derzeit touristisch verstärkt erschlossen werden, könnten einige Populationen durch Bebauung bedroht sein. Über Handel und Wildentnahme ist laut CITES (2005) nichts bekannt.
In der Roten Liste (IUCN, 2004) als uneinstufbar (DD, Data Deficient) angegeben. Nur wenige Informationen über Verbreitung und Populationsgröße liegen vor. Ein Populationstrend ist derzeit nicht einschätzbar. Da die Inseln des Bocas Archipels derzeit touristisch verstärkt erschlossen werden, könnten einige Populationen durch Bebauung bedroht sein. Über Handel und Wildentnahme ist laut CITES (2005) nichts bekannt.
| Beschreibung | Einklappen |
| Größe Sehr kleine Ranitomeya-Art mit einer von 13 - 16 mm. Laut JUNGFER ET AL. (2000) die kleinste bisher bekannte Pfeilgiftfrosch-Art. |
Körper schlank und gestreckt mit ovaler Form. Kopf aber breiter als der Körper. Zähne fehlend. Schnauze von oben betrachtet gerundet und im Profil stumpf gerundet. rund und ca. 40% des Augendurchmessers. Erster Finger deutlich kürzer als der zweite. Rel. Fingerlänge (): 3>4>2>1. Finger mit verbreiterten Haftscheiben (bei Finger I kaum verbreitert). Keine Spannhäute oder Säume. Rel. Zehenlänge (): 4>3>5>2>1. Zehen ohne verbreiterte Haftscheiben. Haut schwach granuliert mit schwarzer Farbe. Darauf zweigt je Seite ein weißlich bis goldener von der Schnauzenspitze ab. Ein in hellweiß bis blau ist vorhanden. R. claudiae besitzt auf der Oberlippe einen in gleicher Farbe. Auf dem Oberarm verläuft eine helle Linie (meist wasserblau) die sich zum Schenkelansatz verbreitert und einen so genannten Signalfleck bildet, welcher heller gefärbt ist als der Rest der Linie. Eine solche Linie findet sich auch auf dem Oberschenkel, dehnt sich dort aber auch oft zu einer unregelmäßigen Fleckung aus. Eine dorsale kann vorhanden sein, ist aber oft unterbrochen. Der Bauch ist weißlich bis hellblau mit schwarzer Marmorierung oder Fleckung. (Alle Angaben nach JUNGFER ET AL., 2001 und OSTROWSKI, 2003.)
 Ranitomeya ()claudiae"Buena Esperanza"© 2008 () | Dorsal- und Ventralbild vergleichen: |
Variation:
Einige Inseln des Bocas Archipels beherbergen ihre eigene Variante von R. claudiae. Diese unterscheiden sich im Farbton der Dorsolateralstreifen und in der Färbung der Schenkeloberseiten. Einige Populatioenen besitzen einen stärker ausgeprägten Medianstreifen (Isla Colón). Die Unterschiede in Zeichnung und Farbe sind jedoch nur gering und nicht so ausgeprägt wie z.B. bei verschiedenen Populationen von Oophaga pumilio oder Dendrobates auratus (OSTROWSKI, 2003).
Interessant erscheint die Tatsache, dass jede Inselvariante in ihrer Färbung den jeweils vorkommenden Populationen von Phyllobates lugubris stark ähneln (s. Abb.: 1 - 6) OSTROWSKI (2003) vermutet, dass es sich dabei möglicherweise um eine Müllersche handeln könnte. Ein Phänomen welches für R. imitator und R. variabilis bereits untersucht wurde und für diese beiden Arten als wahrscheinlich gilt (SYMULA ET AL., 2001).
Interessant erscheint die Tatsache, dass jede Inselvariante in ihrer Färbung den jeweils vorkommenden Populationen von Phyllobates lugubris stark ähneln (s. Abb.: 1 - 6) OSTROWSKI (2003) vermutet, dass es sich dabei möglicherweise um eine Müllersche handeln könnte. Ein Phänomen welches für R. imitator und R. variabilis bereits untersucht wurde und für diese beiden Arten als wahrscheinlich gilt (SYMULA ET AL., 2001).
 Abb.1: Ranitomeya claudiae. Variante "Isla Colón". © 2003 Jeff Mettes. |
 Abb.2: Phyllobates lugubris. Variante "Isla Colón". © 2003 Jeff Mettes. |
 Abb.3: Ranitomeya claudiae Variante "Cerro Brucho". © 2003 Jeff Mettes. |
 Abb.4: Phyllobates lugubris. Variante "Cerro Brucho". © 2003 Jeff Mettes. |
 Abb.5: Ranitomeya claudiae. Varinte "Isla Bastimentos". © 2003 Jeff Mettes. |
 Abb.6: Phyllobates lugubris. Varinte "Isla Bastimentos". © 2003 Jeff Mettes. |
Tab.: Namen, Größenangaben und Farbmusterverteilung verschiedener Populationen von Andinobates claudiae
| Farbmorphe | Foto | Alternativ Namen | Verbreitung | Grundfarbe | Zeichnung | Größe [mm] |
| Buena Esperanza |  | |||||
| Cerro Brucho | ||||||
| Colón | ||||||
| Holotyp | ||||||
| Isla Bastimentos |
Bisher nicht bekannt aber wohl ähnlich der verwandten Arten R. fulgurita und R. minuta bis zu 6 Jahren.
Geschlechtsreife:
Bisher liegen keine Haltungsberichte vor. Die nah verwandte Art R. fulgurita ereicht nach etwa 9 Monaten die Geschlechtsreife (JUNGFER ET AL., 1996).
Hautgifte:
Nach SAPORITO ET AL. (2004) sind sind e (PTX: 251D,281A, 223A) und e (aPTX: 267A , 225A, 297A) Hauptkomponenten des Hautgiftes von R. claudiae.
| Gelege und Larven | Einklappen |
Merkmale Gelege:
vergleichen mit:
Entwicklungszeiten:
Die Eier eines beobachteten Geleges hatten einen Durchmesser von 1,5 mm.
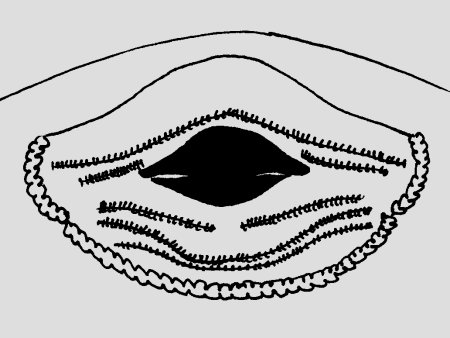 Abb.: ()Mundfeld Ranitomeya claudiae © 2007 () Mundfeld vergleichen: |  Abb.: ()Larve Ranitomeya claudiae © 2007 () |
Entwicklungszeiten:
Ein beobachtetes Gelege benötigte 14 Tage zur Entwicklung (Tagestemperatur: 26 - 28°C). Am 14 Tag waren die Larven verschwunden und wahrscheinlich vom Männchen abtransportiert.
| Verhalten (Ethologie) | Einklappen |
Allgemeines Verhalten:
Revierverhalten:
Balzverhalten:
Brutpflegeverhalten:
Lautäußerung (Vokalisation):
Ruf von Ranitomeya claudiae, © 2005 T. Ostrowski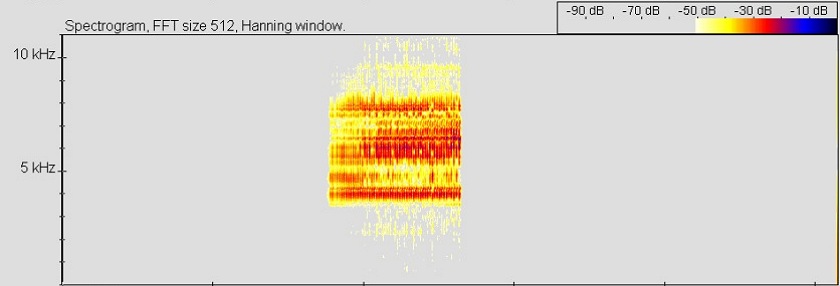
Sonagramm vergleichen:
Ranitomeya claudiae ist ein tagaktiver Bewohner der Laubschicht. Die Tiere leben etwas versteckt sind aber nicht besonders scheu. R. claudiae sind in den Morgenstunden bis etwa 12.00 Uhr besonders aktiv. Zu dieser Zeit lassen sich verstärkt rufende Männchen und Tiere bei der Futteraufnahme beobachten. Gegen Mittag lassen sich dann weniger R. claudiae nachweisen und auch deren Rufaktivität lässt nach. Eine weitere Aktivitätsphase liegt von 15.00 Uhr bis Sonnenuntergang.
Revierverhalten:
Es liegen keine detaillierten Berichte über Populationsstruktur und soziale Interaktion vor. Die Populationsdichte war mit 2-3 Tieren pro qm teilweise sehr hoch (OSTROWSKI, 2003). Der Abstand zwischen rufende Männchen (Revierbesitz?) betrug jedoch meist mehrere Meter. Rufende Männchen fanden sich häufig zwischen alten vertrockneten Blattscheiden am Fuß von Heliconienstauden. Evtl. werden diese gegenüber anderen Männchen als Ablaichplätze oder Larvenabsatzstellen verteidigt. Aggresive Interaktionen konnten aber bisher nicht beobachtet werden (OSTROWSKI, pers. Beobacht.)
Balzverhalten:
Ein von OSTROWSKI (pers. Beobachtung) auf der Isla Colón bei der Balz beobachtetes Paar, zeigte das für Arten der Unterfamilie Dendrobatinae typische Ablaichverhalten. Das Männchen lockte ein Weibchen durch seine zirpenden Rufe an. Dieses näherte sich zielstrebig dem Männchen und begann mit den Vorderbeinen den Hinterleib und Rücken des Männchens einige Male zu berühren (Vorderbeistreicheln). Daraufhin lockte das Männchen das Weibchen zu einem Eiablageort unter einem Blatt. Dies erfolgte unter ständigem Rufen und kurzfristigem Innehalten. Das Weibchen folgte in geringem Abstand und begann beim wiederholten Erreichen des Männchens erneut damit, mit den Vorderbeinen das Männchens zu berühren. Nach etwa 25 min verschwanden die Tiere unter einem Blatt. Die Eiablage wurde nicht beobachtet aber am nächsten Tag konnte an gleicher Stelle ein Gelege nachgewiesen werden. Das Balzverhalten von R. claudiae scheint damit weitgehend dem von R. fulgurita zu entsprechen (JUNGFER ET AL., 1996).
Brutpflegeverhalten:
Das Männchen übernimmt nach dem Laichen die Brutpflege alleine. Ein Männchen konnte auf der Isla Colón einige Tage bei seinem Gelege beobachtet werden. Bei Anwesenheit am Gelege nahm es eine Stellung direkt innerhalb der Galerte ein (s. Abb.) Vermutlich diente diese Körperhaltung der Bewässerung des Geleges. Das Gelege entwickelte sich innerhalb der nächsten 14 Tage vollständig. Innerhalb dieser Zeit wurde das Männchen 5 Mal auf den Eiern angetroffen. Der Schlupf der Larven und deren Abtransport konnten nicht beobachtet werden. Es konnten aber mehrmals Larven tragende Männchen im Biotop beobachtet werden. Absatzstellen der Larven wurden jedoch bisher nicht nachgewiesen. Larven tragende Männchen wurden jedoch auffallend oft im Bereich von Heliconien beobachtet (OSTROWSKI, pers. Beobachtung).
 Abb.: Ranitomeya claudiae beim Bewässern des Geleges © 2003 Thomas Ostrowski |
 Abb.: Das Männchen von Ranitomeya claudiae transportiert die Larven zum Brutgewässer © 2007 Thomas Ostrowski |
 Abb.: Phytothelmata in abgebrochenen Internodien des Riesenbambus. Larvengewässer von R. claudiae.© 2007 Thomas Ostrowski© |
 Abb.: Jungtier und Larve von Ranitomeya claudiae in einer Phytothelmata in einer Bambusinternodie. © 2007 Thomas Ostrowski |
 Abb.: Frisch metramorphisiertes Jungtier von Ranitomeya claudiae. © 2007 Thomas Ostrowski© |
 Abb.: Larve von Ranitomeya claudiae © 2007 Thomas Ostrowski |
Lautäußerung (Vokalisation):
Die Tiere rufen den ganzen Tag über meist versteckt unter Blättern oder in kleinen Höhlen. Der Ruf ist ein leises Insektenartiges Zirpen, welches leicht mit dem Zirpen der vorkommenden tropischen Grillen zu verwechseln ist. Obwohl der Ruf zur typischen Geräuschkulisse im Biotop gehört wurde er erst spät den Tieren zugeordnet, da rufende Männchen nur schwer zu beobachten sind und bei Annäherung das Rufen meist sofort einstellen.
Bei dem Ruf von R. claudiae handelt es sich um einen (chirp-call), der stark den Lautäußerungen der nah verwandten Arten R. minuta und R. fulgurita ähnelt. Fünf von OSTROWSKI bei 25° C aufgenommene und vermessene Rufe der "Colón" - Population von R. claudiae bestanden aus einzelnen Notengruppen mit einer Länge von 879 - 1105 ms und einer Dominanzfrequenz von 5,78 – 5,87 kHz. Jede Notengruppe enthielt 54 – 68 gepulste Noten (Rufrate: 61 – 62 Noten/s). Die Notenlänge betrug im Mittel 15,67 ms und die Noten wurden von Intervallen mit einer Dauer von etwa 1,5 ms getrennt. Anzahl, Länge und Intervall der Pulse konnten mit dem benutzten Programm (BatsoundPro) nicht aufgelöst werden (für weitere Parameter siehe gesamtes Sonagramm).
Ruf von Ranitomeya claudiae, © 2005 T. Ostrowski
Sonagramm vergleichen:
| Lebensraum | Einklappen |
Typenfundort der Erstbeschreibung
Daten von OpenStreetMap - Veröffentlicht unter CC-BY-SA 2.0
Biotop:
Klima:
''Panamá: Pronvincia Bocas del Toro: Festland westlich der Insel Loma Partida (82˚ 11´ W/09˚ 09´ N).'' (JUNGFER ET AL., 2000)
Verbreitung: | 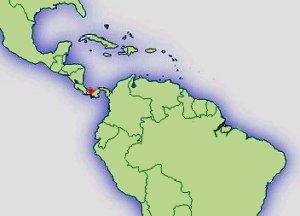 Abb.: ()Verbreitungsgebiet Ranitomeya claudiae © 2008 () |
Biotop:
Die Art scheint kein guter Kulturfolger wie Oophaga pumilio zu sein. Es handelt sich bei allen vom Autor besuchten Biotopen um schattigen Primärwald oder um sehr alte meist verwilderte Kakaoplantagen. Die Biotope lagen fast alle küstennah. Es war meist sehr schattig, und der Boden war mit einer dicken Laubschicht bedeckt. Unterwuchs in Form von Heliconia sp. und Dieffenbachia sp. war meist vorhanden aber nicht besonders dicht. Die bevorzugten Biotope lagen auffallend häufig am Hang und nie in Senken. Diese Beobachtungen decken sich weitgehend mit den Angaben von JUNGFER ET AL. (2000). Die Tiere leben nur bodennah im Laub und konnten nie kletternd beobachtet werden. Teilweise werden alte bewachsene Korallenschutt-Hügel als Biotop genutzt, deren Höhlen und Spalten Versteckmöglichkeiten bieten. Mit D. claudiae waren je nach Fundort meist weitere Dendrobatidae vergesellschaftet (OSTROWSKI, 2003). Es fanden sich häufig Oophaga pumilio, Dendrobates auratus, Phyllobates lugubris, Allobates talamancae, Silverstoneia aff. nubicola und S. cf. flotator im gleichen Biotop. Auf dem Festland bei Cerro Brucho nahe dem Typenfundort kamen fünf! weitere Pfeilgiftfroscharten sympatrisch mit R. claudiae im gleichen vor (OSTROWSKI, pers. Beobachtung). Diese Artenvielfalt wird durch die Besetzung verschiedener ökologischer Nischen möglich. Jede Art nutzt andere Ressourcen im Hinblick auf Aufenthaltsort, Futterangebot und Brutmöglichkeiten.
 Abb.: Laubschicht am Waldboden eines küstennahen Tiefland- regenwaldes auf der Isla Colón. Biotop von R. claudiae.Panama, Provinz Bocas del Toro. © Thomas Ostrowski 2004 |
 Abb.: Wassergefüllte Blätter, hier von Kakao (Theobroma cacao), kommen als Absatzstellen für Larven in Frage.Panama, Provinz Bocas del Toro. © Thomas Ostrowski 2004 |
 Abb.: In der Nähe solcher Heliconienstauden wurden oft rufende oder Larven tragende Männchen beocbachtet.Panama, Provinz Bocas del Toro. © Thomas Ostrowski 2004 |
 Abb.: Wassergefüllten Blattscheiden von Heliconia ssp. könnten ähnlich wie bei R. imitator als Larvenabsatzstellen dienen. Panama, Provinz Bocas del Toro. © Thomas Ostrowski 2004 |
 Abb.: Alte schattige Kakaoplantage. Panama, Provinzia Bocas del Toro. © Jeff Mettes 2003 |
 Abb.: Krautschicht am Waldboden im Primärwald. Panama, Provinzia Bocas del Toro. © 2003 Jeff Mettes |
Klima:
Das südliche Ende des Isthmus (Costa Rica und Panama) wird klimatisch durch die Kordilleren in eine feuchte karibische und eine trockenere pazifische Seite getrennt. Die karibische Seite Panamas zeichnet sich durch ein recht gleichmäßiges tropisches Klima aus. Im Jahresverlauf sind die Temperaturschwankungen sehr gering und betragen im Mittel ca. 1-2°. Auch im Tagesverlauf lassen sich im karibischen Tiefland nur selten höhere Schwankungen zwischen Tag und Nacht nachweisen (WALTER & BRECKLE, 1999). Dennoch kann es in klaren Nächten, nach starken Regenfällen oder bei atlantischen kalten Luftströmungen aufgrund von Hurrikanen in der karibischen See zu kurzfristigen Temperaturstürzen kommen. Im Herbst 2004 kam es in Changuinola, Panama auf Meeresniveau zu einem Kälteeinbruch mit Temperaturen von nur 15 ° C. Auch in Costa Rica konnte der Autor nach starken Regenfällen im Tiefland Temperaturen von 16 ° C messen (OSTROWSKI, pers. Beobachtg.). Diese ungewöhnlichen Tiefsttemperaturen halten allerdings nur wenige Stunden (in Ausnahmefällen einige Tage) an und sind meist nur lokal begrenzt. Auf dem geschützten Waldboden lassen sich aber fast ganzjährig Temperaturen messen, die sowohl am Tag, als auch in der Nacht, etwa dem Jahresmittelwert entsprechen (WALTER & BRECKLE, 2000). Im Gebiet der Provinz Bocas del Toro beträgt diese etwa 26 ° C (s. Abb.). Die Niederschläge sind ebenfalls relativ konstant und ganzjährig hoch. Während des Nordwinters bringen die Luftströmungen des Nordost-Passates feuchte Luftmassen, während im Nordsommer Tiefdruckgebiete über der Karibik für Niederschläge sorgen. Die Niederschläge liegen je nach Abschattung durch die Gebirgszüge zwischen 2000 und 4000 mm im Jahr (WALTER & BRECKLE, 1999). Lediglich in den Monaten Januar bis März kann es lokal zu kurzen Trockenperioden mit weniger als 100 mm im Monat kommen (s. Abb.). Insgesamt sind die Niederschläge über das Jahr aber relativ konstant verteilt, so dass sich die Frösche der karibischen Seite auch das ganze Jahr über fortpflanzen können.
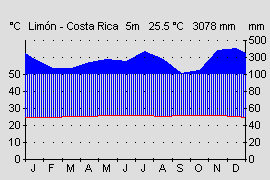 Abb.: Klimadiagramm der Karibikseite in Costa Rica. Gleichmäßige Niederschlagsverteilung ohne Trockenperiode. Quelle: Instituto Geografico Nacional de Costa Rica. | 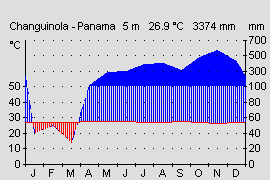 Abb.: Klimadiagramm der Karibikseite in Westpanama. Trockenperiode von Januar bis März. Quelle: www.top-wetter.de |
| Haltung im Terrarium | Einklappen |
Terrarium/Einrichtung:
Temperaturen:
Luftfeuchte:
Ernährung:
Besatz:
Tipps zur Zucht:
Varianten in Terrarienhaltung:
Die Art dürfte zur Zeit nicht legal in Haltung sein. Eine Unterbringung in nicht zu großen Behältern (Futterdichte) von 30 x 30 x 30 cm, wie bei R. fulgurita und R. minuta, sollte für die Haltung eines Paares genügen. Ein kleiner Wasserteil und eine dichte Bepflanzung sowie einige Höhlen () sollten vorhanden sein. Eine Laubschicht aus Buchen- oder Eichenlaub gibt den Tieren Deckung und Ablaichmöglichkeiten.
Temperaturen:
Die Temperatur sollte 28 °C nicht übersteigen, da die Tiere im Schatten in der Laubschicht leben und dort nie mehr als 26 °C nachgewiesen werden konnten. 24-26 °C erscheinen deshalb das Optimum zu sein.
Luftfeuchte:
Die Luftfeuchtigkeit sollte recht hoch liegen (min. 80%) dennoch sollten trockene Stellen vorhanden sein und der Boden nicht zu nass sein.
Ernährung:
Aufgrund der gefundenen Gifte, kann wohl darauf geschlossen werden, dass das Beutespektrum der Art in der Natur ebenfalls wie bei O. pumilio oder D. auratus zum großen Teil aus Ameisen besteht. Bestimmte Ameisenarten enthalten nämlich die in der Froschhaut angereicherten e (SAPRITO ET AL., 2004) und machen z.B. bei O. pumilio etwa 80% der Nahrung aus (DONNELLY, 1991). R. claudiae bewältigt sicher nur kleinste Ameisenarten, da er nach JUNGFER ET AL. (2000) wohl der bisher kleinste Pfeilgiftfrosch überhaupt ist. Im Terrarium kommen deshalb nur Futtertiere in Springschwanzgröße für eine Ernährung in Frage. Schon kleine Drosophila dürfte die Art nur noch schwer überwältigen können. Eine Haltung ist nur mit gut laufender Springschwanzzucht Erfolg versprechend und für die Aufzucht der Jungtiere sicher unerlässlich.
Besatz:
Es sollten in den kleinen Becken nur ein Paar oder ein Männchen mit zwei Weibchen gehalten werden. In größeren Becken evtl. mehr, allerdings könnte dann die Futterversorgung schwierig werden. Im Biotop lag die Besatzdichte bei 2-3 Tieren pro Quadratmeter (OSTROWSKI, 2003) allerdings hatten rufende (Revier besitzende) Männchen einen etwas größeren Abstand von 2-3 m zueinander (OSTROWSKI, pers. Beobachtg.).
Tipps zur Zucht:
Bisher nicht nachgezüchtet. Wie bei den sehr nah verwandten Arten R. fulgurita und R. minuta, wird die Eiablage auf Blättern oder in kleinen Höhlen (Fotodosen) auf waagerechten glatten Flächen erfolgen. Die Larven dürften, wie die der verwandten Arten, ebenfalls sein und sich gut mit einem Gemisch aus verschiedenen Fischfuttersorten aufziehen lassen.
Varianten in Terrarienhaltung:
| Buena Esperanza |  |
| Cerro Brucho |  |
| Colón |  |
| Holotyp |  |
| Isla Bastimentos |  |
Daten errechnet aus von Nutzern gemeldeten Fröschen (Home->Meine Frösche) und den aktuellen Inseraten.
stand 18.06.2026 02:14 Uhr
stand 18.06.2026 02:14 Uhr
| Autoren dieser Seite: | Einklappen |
| Referenzen | Einklappen |
BIOPAT (2005): Patenschaft für biologische Vielfalt. - www.biopat.de (Stand: 27.12.2005)
CITES (2005): Convention on International Trade of Endangered Species of Wild Fauna and Flora.
XXOnlineversion: 11.08.05. Electronic Database accessible at http://www.cites.org/index.html
DONNELLY, M.A. (1991): Feeding Patterns of the Strawberry Poison Frog, Dendrobates pumilo,
XX(Anura: Dendrobatidae). - Copeia 1991: 723-730
FROST, D.R. (2007): Amphibian Species of the World: an Online Reference. Version 5.0 (1.02.2007).
XXElectronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.php
XX Am.Mus.Nat.Hist. New York, USA.
GRANT, T . , FROST, D. R. , CALDWELL, J. P. , GAGLIARDO, R. , HADDAD, C. F. B. , KOK, P. J. R. ,
XXMEANS, D.B. , NOONAN, B. P. , SCHARGEL, W. E. & W.C. WHEELER (2006): Phylogenetic
XXsystematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae).
XX - Bulletin of the Am.Mus.Nat.Hist. 299. (PDF)
IUCN, Conservation International, and NatureServe. (2009): Global Amphibian Assessment.
XXwww.globalamphibians.org. Downloaded on 15 October 2004.
JUNGFER, K.-H., H. BIRKHAHN, V. KÜLPMANN & K. WASSMANN (1996): Haltung und Fortpflanzung von
XXDendrobates fulguritus SILVERSTONE, 1975, mit Anmerkungen zur Gattung Minyobates MYERS, 1987.
XX- Herpetofauna. 18(104):19-27.
JUNGFER, K.-H., S. LÖTTERS & JÖRGENS, D. (2000): Der kleinste Pfeilgiftfrosch - eine neue
XXDendrobates-Art aus West-Panama. - Herpetofauna 22 (129): 11-18.
METTES, J. (2003): A Lesser-known Panamanian Dendrobatid.
XX- http://www.geocities.com/danstukky/claudiae.html (Stand: 23.05.2003)
OSTROWSKI, T. (2003): Dendrobates claudiae und Phyllobates lugubris - zwei Pfeilgiftfrösche aus
XXPanama im gleichen Kleid. - Reptilia 8 (5): 72-75 (PDF)
SAPORITO, R.A., GARRAFFO, H.M., DONNELLY, M.A. , EDWARDS, A.L., LONGINO, J.T., & J.W. DALY (2004):
XXFormicine ants: An arthropod source for the pumiliotoxin alkaloids of dendrobatid poison frogs.
XX- Proc.Nat.Acad.Sci USA 101 (21):8045-8050
SYMULA, R., SCHULTE, R. & K. SUMMERS (2001): Molecular phylogenetic evidence for a mimetic
XXradiation in Peruvian poison frogs supports a Müllerian mimicry hypothesis.
XX- Proc.R.Soc.Lond.B. 268 (1484): 2415 - 2421
WALTER, H. & S. BRECKLE (1999): Vegetationen und Klimazonen. 7. Auflage. - Ulmer, Stuttgart.
CITES (2005): Convention on International Trade of Endangered Species of Wild Fauna and Flora.
XXOnlineversion: 11.08.05. Electronic Database accessible at http://www.cites.org/index.html
DONNELLY, M.A. (1991): Feeding Patterns of the Strawberry Poison Frog, Dendrobates pumilo,
XX(Anura: Dendrobatidae). - Copeia 1991: 723-730
FROST, D.R. (2007): Amphibian Species of the World: an Online Reference. Version 5.0 (1.02.2007).
XXElectronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.php
XX Am.Mus.Nat.Hist. New York, USA.
GRANT, T . , FROST, D. R. , CALDWELL, J. P. , GAGLIARDO, R. , HADDAD, C. F. B. , KOK, P. J. R. ,
XXMEANS, D.B. , NOONAN, B. P. , SCHARGEL, W. E. & W.C. WHEELER (2006): Phylogenetic
XXsystematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae).
XX - Bulletin of the Am.Mus.Nat.Hist. 299. (PDF)
IUCN, Conservation International, and NatureServe. (2009): Global Amphibian Assessment.
XXwww.globalamphibians.org. Downloaded on 15 October 2004.
JUNGFER, K.-H., H. BIRKHAHN, V. KÜLPMANN & K. WASSMANN (1996): Haltung und Fortpflanzung von
XXDendrobates fulguritus SILVERSTONE, 1975, mit Anmerkungen zur Gattung Minyobates MYERS, 1987.
XX- Herpetofauna. 18(104):19-27.
JUNGFER, K.-H., S. LÖTTERS & JÖRGENS, D. (2000): Der kleinste Pfeilgiftfrosch - eine neue
XXDendrobates-Art aus West-Panama. - Herpetofauna 22 (129): 11-18.
METTES, J. (2003): A Lesser-known Panamanian Dendrobatid.
XX- http://www.geocities.com/danstukky/claudiae.html (Stand: 23.05.2003)
OSTROWSKI, T. (2003): Dendrobates claudiae und Phyllobates lugubris - zwei Pfeilgiftfrösche aus
XXPanama im gleichen Kleid. - Reptilia 8 (5): 72-75 (PDF)
SAPORITO, R.A., GARRAFFO, H.M., DONNELLY, M.A. , EDWARDS, A.L., LONGINO, J.T., & J.W. DALY (2004):
XXFormicine ants: An arthropod source for the pumiliotoxin alkaloids of dendrobatid poison frogs.
XX- Proc.Nat.Acad.Sci USA 101 (21):8045-8050
SYMULA, R., SCHULTE, R. & K. SUMMERS (2001): Molecular phylogenetic evidence for a mimetic
XXradiation in Peruvian poison frogs supports a Müllerian mimicry hypothesis.
XX- Proc.R.Soc.Lond.B. 268 (1484): 2415 - 2421
WALTER, H. & S. BRECKLE (1999): Vegetationen und Klimazonen. 7. Auflage. - Ulmer, Stuttgart.
| Zitierung: | Einklappen |
OSTROWSKI, T. & T. MAHN (2026): Artbeschreibung Andinobates claudiae.
- Dendrobase.de - Eine Online-Datenbank der Familie Dendrobatidae (Anura).
https://www.dendrobase.de/html/D_andinobates_claudiae.html; Stand: Juni 2026
- Dendrobase.de - Eine Online-Datenbank der Familie Dendrobatidae (Anura).
https://www.dendrobase.de/html/D_andinobates_claudiae.html; Stand: Juni 2026
| Link für Website Betreiber: | Einklappen |
Webmaster dürfen Links von Ihren Seiten zur DendroBase erstellen, wenn sie folgende Kriterien erfüllen:
- Die Website enthält weder illegale, rassistische oder sexistische Inhalte
- Das Thema der Website bezieht sich auf Frösche
- Der Link genau wie folgend angegeben verwendet wird:
<a href="https://www.dendrobase.de/html/D_andinobates_claudiae.html" target="_new">Andinobates claudiae (DendroBase.de)</a>
- Die Website enthält weder illegale, rassistische oder sexistische Inhalte
- Das Thema der Website bezieht sich auf Frösche
- Der Link genau wie folgend angegeben verwendet wird:
<a href="https://www.dendrobase.de/html/D_andinobates_claudiae.html" target="_new">Andinobates claudiae (DendroBase.de)</a>
| Fotos | Einklappen |

Ranitomeya ()claudiae
© 2003 ()

Ranitomeya ()claudiae "Colón"
© 2003 ()

Ranitomeya ()claudiae "Colón"
© 2003 ()

Ranitomeya ()claudiae
"Cerro Brucho"
© 2003 ()

Ranitomeya ()claudiae "Colón"
© 2004 ()

Ranitomeya ()claudiae "Colón"
© 2004 ()

Ranitomeya ()claudiae
"Buena Esperanza"
© 2008 ()

Ranitomeya ()claudiae
"Buena Esperanza"
© 2008 ()
DendroBase.de Version 5 | Impressum | Datenschutzerklärung